

生殖隔离指由于各方面的原因,使亲缘关系接近的类群之间在自然条件下不交配,或者即使能交配也不能产生后代或不能产生可育性后代的隔离机制,若隔离发生在受精以前,就称为受精前的生殖隔离。
基本信息对真核生物来说, 无论它们在形态上的差别有多大, 生殖隔离(reproductive isolation)应该是两个群体能否真正分化成不同物种的关键, 这种隔离机制可以是地理的、行为的或其他方式; 而生殖隔离总会伴随着一些形态或遗传上的变化, 虽然这些特征可能与生殖隔离本身并无多大关系, 但往往成为分类学家或分子进化生物学家区分种的依据。
生殖隔离大致相当于从前所说的生理隔离,与地理隔离既有联系,也有区别。地理隔离由外因引起,而生殖隔离一般是在具有遗传差异(内因)的基础上才有可能(遗传隔离)。同一地点的种、亚种的隔离完全是生殖隔离,但不同地点的种、亚种在许多情况下,似乎也会很快发生这种作用。关于生殖隔离,许多人进行过讨论。斯特宾斯(G.L.Stebbins,1966)作了如下的分类:(1)合子形成前的机制。(i)生态隔离:虽在同一地域,但生态地位不同。(ii)季节隔离:开花期有分离。(iii)行为性隔离(ethological isola-tion):雌雄的性习性不同。(iv)机械隔离:基于生殖器官构造的差异。(v)配子不亲和现象(gameteincompatibility):一方的配子体在另一方的生殖器官内死亡。(2)合子形成后的机制。(i)杂种死灭或弱势(hybrid inviability,hybrid weakness)。(ii)杂种不育。(iii)杂种衰败(hybrid break-down):F2以后死灭或弱势。在自然界中这些因子是重复地起着隔离作用的。
 因生殖方面的原因使亲缘关系相近的类群间不杂交,或杂交不育,或杂交后产生不育性后代的现象。实质是不同种群内的个体,因受不同基因型控制而有不同的生殖行为及相关生殖率,故种群间的基因流动受到限制或被阻止。根据影响基因流动的障碍不同分为:①外在生殖隔离。基因流动的障碍是个体受精前外在的影响,包括机械隔离、行为隔离、季节隔离和配子隔离。如菊科莴苣属(Latuca)的L. canadensis和L. gra- minifolia,分布于美国东南部,人工杂交可育,但在自然界中,前者夏季开花而后者春季开花,得以保持两个个别的形态各异的生殖隔离种;②内在生殖隔离。基因流动受到个体内部的影响。由不同群体的配子体、配子、染色体或基因间不能相互作用所致,包括不亲和性、杂种不存活、杂种不孕、杂种破落等4种隔离。如马(Equus caballus)和驴(E. asinus)杂交,不能产可育的杂种后代。地理隔离和生态隔离都含有生殖隔离的意义。生殖隔离是隔离的重要内容,是物种演化的一个因素和物种形成的必要条件。在成种过程中,一般先有地理隔离,后有生殖隔离。1
因生殖方面的原因使亲缘关系相近的类群间不杂交,或杂交不育,或杂交后产生不育性后代的现象。实质是不同种群内的个体,因受不同基因型控制而有不同的生殖行为及相关生殖率,故种群间的基因流动受到限制或被阻止。根据影响基因流动的障碍不同分为:①外在生殖隔离。基因流动的障碍是个体受精前外在的影响,包括机械隔离、行为隔离、季节隔离和配子隔离。如菊科莴苣属(Latuca)的L. canadensis和L. gra- minifolia,分布于美国东南部,人工杂交可育,但在自然界中,前者夏季开花而后者春季开花,得以保持两个个别的形态各异的生殖隔离种;②内在生殖隔离。基因流动受到个体内部的影响。由不同群体的配子体、配子、染色体或基因间不能相互作用所致,包括不亲和性、杂种不存活、杂种不孕、杂种破落等4种隔离。如马(Equus caballus)和驴(E. asinus)杂交,不能产可育的杂种后代。地理隔离和生态隔离都含有生殖隔离的意义。生殖隔离是隔离的重要内容,是物种演化的一个因素和物种形成的必要条件。在成种过程中,一般先有地理隔离,后有生殖隔离。1
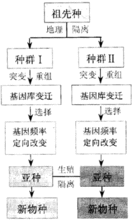
分析现代进化论认为,物种形成大致要经过3个阶段:1)由于基因突变、染色体变异和基因重组,使种群中产生可遗传的变异;2)自然选择等因素作用于可遗传的变异,使种群的遗传结构(基因型频率和基因频率)发生了适应性的改变;3)不同种群由于地理隔离和生态隔离而加深了性状分歧,逐渐形成亚种,一旦出现了生殖隔离,亚种就变成了新种。此外,在植物界异源多倍体的产生亦可导致新种形成。2
分类地理隔离同一种生物由于地理上的障碍而分成不同的种群,使种群间不能发生基因交流的现象,叫做地理隔离。1
生态隔离由于食性、生活习性和栖息地点的不同,使几个亲缘关系接近的类群之间交配不易成功的隔离机制。例如有人报道5种近缘雀在树冠的不同层次捕食不同的昆虫,以及营巢的位置和交配产卵的繁殖季节不同,所以它们虽然生活在同一片森林,甚至同一棵树上,也都因各有自己的生态位置而形成生殖隔离。
季节隔离又称时间隔离,新种形成过程中,合子形成前的隔离机制之一。生物一般都有一定的生育季节和时间,如动物的发情期、交配期;植物的开花期和授粉期等,如果同种群体间的生育季节和时间不同,就会造成季节隔离或时间隔离,阻止了基因的交流,从而导致生殖隔离的形成。例如,北美有3种蛙,同生在一个池塘里,但由于繁殖季节不同而构成了生殖隔离。林蛙在水温达到7 ℃时,首先产卵;其次是笛蛙,在水温达到12 ℃时开始产卵;鼓蛙在水温达到16 ℃时才开始产卵。
生理隔离植物传粉后或动物交配后,由于生理上的不协调而不能完成受精作用的现象。例如,异种花粉落在柱头上通常不能形成花粉管;有的即使形成花粉管也不能成功地通过花柱、到达胚珠。又如绿果蝇的精子在美洲果蝇的受精囊内很快就丧失游动能力,而同种精子则可较长时间保持活动能力。
机械隔离主要指由于植物花的结构与传粉者形态结构不协调而造成的交配受精障碍。例如,大红吊钟柳,花红色,有细长的管状花冠;山吊钟柳,花蓝色,有较大的钟状花冠。二者在人工授粉时能形成杂种,但在自然条件下,前者以其红色光和红外光招引蜂鸟,管状的花冠对蜂鸟细长的喙也相适应;后者以蓝紫光与紫外光招引蜂类,钟状花冠也适于蜂类采蜜,但二者之间不能相互授粉。这样,它们的花色和结构,便构成了物种间的隔离而阻止了基因的交流,此即为形态隔离。
行为隔离由于交配行为不同,而使两个或几个亲缘关系相近的类群之间交配不易成功的隔离机制。例如,红蟋蟀、宾州蟋蟀和富尔顿蟋蟀在形态上无区别,但鸣声的强弱和频率不同。试验证明,交配仅在发生同一鸣声的种内进行,不同鸣声的蟋蟀不进行交配。
杂种不活一种杂交后的生殖隔离,不同种生物受精后,虽能形成合子,但合子不能存活;或合子虽能存活,但胚胎发育到一定阶段便死亡;或者胚胎发育虽然完成,并发育为幼体,但不能生活到性成熟,故留下后代的概率很低。例如,以奥地利亚麻(♀)与多年生亚麻(♂)杂交,杂种的胚不能充分发育,因为胚和胚乳不协调。反交时,胚和胚乳虽较协调,杂种胚可以充分发育,但不能穿破母本组织形成的种皮,种子不能萌发,杂种也不能成活。
杂种不育一种杂交后的生殖隔离。不同种生物杂交后虽能形成合子,并发育为成熟的F1杂种,但杂种不能产生后代。例如,马和驴杂交所生的骡就不能生育。因为马的染色体为64,驴的染色体数为62,骡的染色体数为63。马的32对染色体中有18对端着丝粒染色体,驴的31对染色体中只有11对端着丝粒染色体,骡在形成生殖细胞时,染色体不能正常配对(即联会紊乱)而不能完成减数分裂和形成正常的配子,故不能生育。3
杂种衰败也叫杂种破落,是一种杂交后的生殖隔离机制。种间杂交虽能形成能育的F1,但在F2或回交后代中,绝大部分重组基因型不相协调,无适应能力,不能存活。例如,有人以陆地棉的斑点变种为母本,与夏威夷棉杂交,F1能活也能育。但是F1植株上结出的106粒F2种子中,7粒胚小,不能发芽;32粒胚看来正常,但也不能萌发;9粒能穿破种皮,但子叶不能展开;22粒能长成幼苗,但不久即死亡;16粒长成弱苗,仅活了3周;20粒长成的幼苗,能活3周以上,但以后发育也不正常。这表明在F2的重组基因型中,存在着杂种衰败机制,它们经不起强大的自然选择压力,最终也不能存活下来。
以上各种类型的生殖隔离,实质上都是阻碍不同物种间的基因交流,使各个隔离种群各有较强的稳定性,以保证在自然条件下各按着与环境相适应的方向发展。因此,物种是生殖隔离的种群,由地理隔离发展到生殖隔离是大多数物种形成的基本因素。2
生殖隔离的遗传机制远缘杂交的实质是打破物种间的生殖隔离,实现不同物种间的优良基因重组,创造为人类所需的植物新类型和新种质。物种间的生殖隔离是由不同的繁殖隔离机制来保持的,繁殖隔离机制阻止种间的基因流动,是防止生物种间杂交的生物学特性。根据隔离发生的主要时期将繁殖隔离机制划分为两大类:受精前的生殖隔离和受精后的生殖隔离。若隔离发生在受精以前,就称为受精前的生殖隔离,其中包括地理隔离、生态隔离、季节隔离、生理隔离、形态隔离和行为隔离等;若隔离发生在受精以后,就称为受精后的生殖隔离,其中包括杂种不活、杂种不育和杂种衰败等。3
水稻生殖隔离机理亚洲栽培稻分为籼稻和粳稻两个亚种,籼粳亚种间的杂种优势要远大于籼粳亚种内的优势。但由于自然界普遍存在的亚种之间生殖隔离法则,籼粳稻亚种间杂种育性下降, 结实率很低。受此制约,长期以来杂交水稻的育种工作基本限制在亚种内进行。然而,到 20 世纪 80 年代水稻科研工作者发现,S5可能是控制水稻籼粳杂种不育性状的主效基因位点。2008 年,张启发课题组与多家研究单位合作成功克隆了 S5 位点的 5 号基因,并发现仅 5 号基因的作用无法解释该位点引起杂种不育的机理。4
本词条内容贡献者为:
胡芳碧 - 副教授 - 西南大学
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国