

物质跨膜运输是细胞维持正常生命活动的基础之一,也是细胞膜的重要功能之一。通过跨膜运输,可以沟通细胞内外及细胞内各细胞器之间的联系,保证新陈代谢等生命活动中的正常物质交换,也是生物膜能量转换和信息传递等功能的基础。
物质跨膜运输的方式可分为两大类,一类是小分子和离子物质的跨膜运输,包括主动运输和被动运输,另一类是大分子和颗粒物质的膜泡运输,包括胞吞作用和胞吐作用。1
主动运输主动运输(activetransport)是通过消耗能量,膜运输蛋白由低浓度侧向高浓度侧逆浓度梯度转运离子或分子的运输方式。根据能量利用方式的不同,主动运输分为ATP驱动泵运输和协同运输两种类型。2
ATP驱动泵ATP驱动的跨膜转运蛋白即ATP驱动泵,具有催化ATP水解的酶活性,可利用ATP水解释放的能量逆浓度梯度跨膜转运离子或分子,共有四种类型,即P类、V类, F类和ABC超家族类。P类、V类及F类都与离子转运有关。Na+-K+ 泵和Ca2+泵属于P类转运蛋白,此类转运蛋白的胞内α亚基可结合并催化ATP水解,使自身发生磷酸化。F类和V类转运蛋白负责转运质子,也称为H泵。F类转运蛋白主要存在于细菌的质膜和真核细胞线粒体内膜上,负责将H+运入线粒体,供ATP合成,也称为ATP合酶。V类转运蛋白负责将过多的H+泵出细胞,以维持细胞内pH值的稳定。ABC超家族转运蛋白主要介导糖类、脂类、氨基酸、无机酸分子,肽类如谷胱甘肽衔生物以及生物体内的次生代谢物和外来毒索的运输。2
Na+-K+泵
是运输Na+和K+离子的跨膜运输蛋白,因具有ATP酶活性,故又称Na+-K*ATP酶。Na+-K+泵由α、 β两个亚单位组成,胞质面有3个Na+和1个ATP结合位点,胞外表面有2个K+结合位点和乌本苷(Na+-K+泵的抑制剂)结合位点。Na+-K+泵每秒可水解1000个ATP分子,每水解1个ATP分子向胞外运出3个Na+,向胞内运进2个K+。
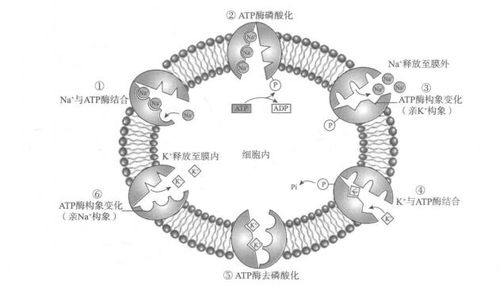
Na+-K+泵的工作原理是:当Mg2+存在时,胞质中的Na+与此酶的Na+离子位点结合,酶的α亚基被激活,消耗ATP发生酶的自身磷酸化,同时ATP水解为ADP;自身磷酸化后酶构象改变,将Na+转运到胞外,此时Na+与变构的α亚基亲和力降低,从酶上释放,处于该构象的酶与膜外侧K+亲和力高,结合K+离子后,刺激酶去磷酸化并恢复原构象,将K+离子转运到胞内并释放,完成一次运输。
细胞消耗大量的ATP来维持细胞内外Na+、 K+离子梯度差,具有重要的生理意义,如调节细胞渗透压、维持膜电位,驱动某些营养物质的主动吸收(如葡萄糖的转运)等。氰化物等有毒物质可使ATP供能中断,造成Na+-K+泵的工作停止。2
Ca泵
又称钙ATP酶,是运输Ca2+离子的跨膜运输蛋白,存在于细胞膜或某些细胞器膜上,具有ATP酶活性,可发生磷酸化与去磷酸化反应,将Ca2+逆浓度泵出细胞或泵入某些细胞器,维持细胞内Ca2+的低浓度水平(≤10-7mol/L),此浓度是细胞外液(约10-3mol/L)的万分之一。如同Na+-K+泵的工作原理一样,Ca2+泵每水解一个ATP分子,通过自身构象变化,将胞质中2个Ca2+逆浓度转运到细胞外或泵入肌质网,通过细胞内Ca2+浓度的变化调节细胞运动、肌细胞收缩、细胞分泌. 神经递质的释放及细胞信号转导等。
除Na+-K+泵,Ca2+泵外,体内还有两种重要的离子泵,一种是位于胃腺壁细胞膜和肾小管细胞膜上的H+-K+ATP酶,主要功能是分泌H+,另一种是分布在细胞器膜上的H+-ATP酶,可将H+由胞质泵入容酶体、胞内体及突触囊泡等结构内,维持细胞器内特定的pH值。2
ATP供能的ABC超家族转运
ABC (ATP-binding cassette)转运蛋白由两个跨摸结构域和两个ATP结合域组成,当ATP结合域与ATP结合后,两个ATP结合域发生二聚化,伴随着ATP水解成ADP,二聚体解离。ABC转运蛋白的工作原理是:胞内疏水性底物分子首先转移到质膜内侧,进入质膜内叶,继而底物分子侧向扩散至ABC转运蛋白并与之结合,ABC转运蛋白将底物分子转运到质膜内叶,脱离膜脂游离到细胞外,完成物质的转运。ABC转运蛋白是一个超家族,已在哺乳类细胞鉴定其家族成员有近50种,在参与药物代谢,物质吸收和代谢废物排泄的肝脏、小肠及肾等组织细胞广“泛存在,主要是介导生物体内的次生代谢物的运输。
多药耐药性(multidrug resistance,MDR)是肿瘤细胞在某种抗肿瘤药物作用下出现耐药性,并对其他多种结构和作用机制不同的抗肿瘤药物也产生交叉耐药性。MDR蛋白是ABC家族中的一员,负责将细胞中亲脂性小分子物贡转运出细胞。临床上使用的抗肿瘤药物多为脂溶性小分子,当癌细胞中MDR蛋白高表达时,MDR蛋白会将进入细胞的亲脂性抗癌药物转运出胞,使抗癌药物失去杀伤癌细胞的作用,故此产生多药耐药性。2
协同运输协同运输(co-transport)是指一种物质顺浓度梯度的被动运输驱动了另一种物质逆浓度梯度进行主动运输的过程。如果两种物质运输方向相同,称为同向运输,如果方向相反,称为对向运输。动物细胞的协同运输通常是由膜两则的Na+电化学梯度来驱动,植物细胞和细菌通常是由H+电化学梯度来驱动。
同向运输
是两种溶质分子向同一方向的穿膜运输。在这种方式中,物质逆浓度梯度穿膜运输方向与其所依赖的离子顺浓度梯度的穿膜运输方向相同。小肠上皮细胞吸收葡萄糖的过程就是与Na+的同向协同运输。在小肠上皮细胞内,葡萄糖浓度比肠腔高,尽管如此,肠腔内的葡萄糖仍逆浓度不断运输进人小肠上皮细胞内。葡街糖逆浓度转运的动力来自小肠上皮细胞内外的Na+离子的势能:Na+-K+ATP 酶不断地向细胞外“泵”Na+,结果是细胞外高浓度的Na+顺电势差通过小肠上皮细胞顶面的Na+载体蛋白再被动回流到小肠上皮细胞内,但Na+载体蛋白结合Na+的同时与葡萄糖偶联,顺势将葡萄糖携带到胞内。在胞内Na+释放后,Na+载体蛋白构象变化,失去与葡萄糖分子的亲和力而与之分离,葡萄糖释放到细胞内。 在小肠上皮细胞的基底面,葡萄糖由其特定的载体蛋白再经易化扩散转运至血液。Na+与葡萄糖的同向运输保证了葡萄糖吸收和运输的方向性。2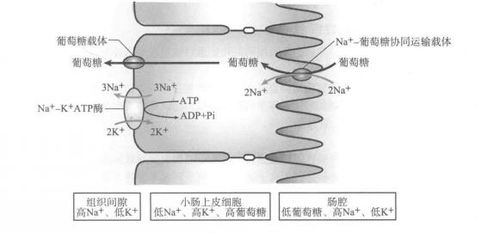
对向运输
是在离子浓度梯度驱动下,由同一种载体蛋白将两种不同的离子或分子分别向膜的相反方向转运的穿膜运输过程。高等动物的细胞都有一种或多种对向运输载体,如Na+-H+交换载体(Na+-H+exchange carrier),它可以通过Na+顺浓度梯度内流把H+泵出细胞,从而清除细胞代谢产生的过量H+,细胞内pH维持在7.2左右。2
被动运输被动运输(passive transport)是物质顺浓度梯度且不消耗细胞代谢能(ATP)所进行的运输方式,运输动力来自质膜内、外侧物质的浓度梯度势能或电位差。被动运输分为简单扩散和易化扩散。2
简单扩散简单扩散(simple diffusion)是脂溶性小分子物质穿膜运输的方式,被运输的小分子物质在膜两侧存在浓度差,即可顺浓度进行扩散。小分子物质扩散的速度取决于分子的大小和其脂溶性,如疏水性气体小分子O2、CO2、N2,等能迅速通过细胞膜。乙醇等脂溶性高的小分子也能迅速通过细胞膜。H2O、尿素、甘油等分子带有强极性,脂溶解度低,故通过膜比较缓慢。更大的亲水性分子如葡物糖和各种带电的分子如氨基酸及核苷酸等都不能经简单扩散的方式进行运输。2
易化扩散易化扩散(facilitated diffusion)又称协助扩散,是借助于膜上运输蛋白的协助顺浓度梯度运输物质的方式。葡萄糖、氨基酸,核苷酸等亲水性营养物质以及Na+,K+,Ca2+等带电荷的离子都能通过易化扩散进行跨膜运输。介导易化扩散的运输蛋白有载体蛋白和通道蛋白,据此,易化扩散又分为载体蛋白介导的易化扩散和通道蛋白介导的易化扩散两种类型。介导主动运输的蛋白也称为载体蛋白。2
载体蛋白介导的易化扩散
运输过程是通过载体蛋白发生可逆的构象变化实现的。载体蛋白是膜上与物质运输有关的穿膜蛋白,对所运输的物质具有高度选择性,当载体蛋白一端表面的特异结合部位与专一的溶质分子结合,引发载体蛋白空间构象改变,将运送的溶质分子从结合的一侧转运到膜的另一侧;变构的载体蛋白对被转运物质的亲和力同时发生改变,于是被转运的溶质分子与载体蛋白分离而被释放,载体蛋白又恢复到原来的构象。载体蛋白通过周而复始的构象变化被反复循环使用。
易化扩散的速率取决于膜两则溶质分子的浓度差。随着浓度差增大,运输速度加快,但当溶质分子与载体蛋白结合的位点饱和时,运输速率达到饱和,不会再增大。载体蛋白的活性可被调节,其中激索起主要调节作用。
大多数哺乳类动物细胞膜上都含有协助葡萄糖从血液扩散到细胞内的葡萄糖转运蛋白(glucose transporter, GLUT), 以易化扩散方式将能萄糖转运人细胞,如人红细胞膜上约有5万个葡萄糖转运蛋白,其数量相当于膜蛋白质总量的5%,最大转运速度约为每秒转运180个葡萄糖分子。葡萄糖转运蛋白家族包括GLUT1-GLUT14,均参与葡萄糖的运输。GLUT1分布于多种细胞的质膜上,对葡萄糖亲和力高,易于葡萄糖吸收进人细胞。GLUT2 主要分布在肝细胞、胰岛β细胞(啮齿类)及小肠和肾具吸收功能的上皮细胞,与葡萄糖的结合具有高泡和度和低亲和性特点。GLUT3分布于脑内神经元细胞,对葡笋糖有很高的亲和力和运输容量,即使在血糖水平略低时也能迅速从细胞外液转运葡萄糖,保障神经元的能量供应。GLUT4分布于肌细胞和脂肪细胞膜上,胰岛索可调节GLUT4的数量。正常情况下,靶细胞的GLUT4以囊泡形式贮存在细胞内,当餐后血糖升高时,刺激胰岛细胞分泌胰岛索,胰岛素刺激靶细胞,使含GLUT4的胞内囊泡迅速移至细胞表面,GLUT4 插入质膜中,以提高葡萄糖的摄取,保证血糖的稳定。相反,当机体处于饥饿状态时,血糖浓度降低,在胰高血糖素作用下,肝糖原降解,产生大量葡萄糖,细胞内葡萄糖糖浓度高于细胞外,葡萄糖结合到载体蛋白细胞内侧的结合部位上,将葡笋唐运到细胞外。糖尿病病人常伴有GLUT4数量不足或功能下降,血糖升高时,葡萄糖不能顺利进人靶细胞,导致血糖持续增高,是产生胰岛索抵抗的原因之一。2
通道蛋白介导的易化扩散
运输过程借助于穿越脂双分子层的通道蛋白完成。通道蛋白中心是亲水性小孔,不同种类的通道蛋白可分别运输离子,水等小分子。主要运输离子的通道蛋白也称为离子通道,对离子具有高度亲和力和高度选择性。离子通道运输速率高,每秒运输离子数量多达几百万个,载体蛋白每秒运载的分子数目则不足一千个。某些离子通道蛋白星关闭状态,在接受特定信号刺激时发生构象的变化而开启,开放时间不超过几毫秒,随即关闭,相当于闸门,也称为闸门离子通道。
根据闸门离子通道开启信号的不同分为3类。第一类是电压门通道,通道的开、闭受膜电位变化控制,如Na+通道、K+通道等。第二类是配体门通道,通道的开、闭受化学物质调节,研究较深入的是乙酰胆碱受体通道。第三类是应力激活通道,通道蛋白感受机械应力而改变构象,开启通道。如内耳听觉毛细胞顶部的听毛就具有这种通道,声波刺激在耳蜗内转换成机械振动,可引起听毛倾斜,牵动听毛上应力活化的K+通道,K+流人内耳毛细胞,使毛细胞发生去极化,产生听觉信号。
闸门离子通道的瞬时开关有利于细胞各种功能活动依次进行。神经冲动引起骨骼肌收缩的整个反应过程至少有4种闸门离子通道被依次激活,顺序开放和关闭。
①当冲动到达神经末梢,神经细胞膜发生去极化,膜电位降低,引起神经末梢膜上的电压闸门Ca2+通道开放,膜外高浓度的Ca2+迅速进人神经末梢,刺激分泌神经递质——乙酰胆碱。
②释放的乙酰胆碱与肌肉细胞膜上的配体闸门通道上的特异部位结合,闸门瞬间开放,Na+大量进入细胞,引起肌细胞膜电位改变,膜局部去极化。
③肌肉细胞膜的局部去极化又使其膜上的电压闸门Na+通道开放,更多的Na+进入肌细胞,去极化扩展到整个肌细胞膜。
④肌细胞膜去极化又引起肌细胞内肌质网上的Ca2+通道开放,从肌质网内流人细胞质,肌细胞内Ca2+浓度急剧升高,引发肌原纤维收缩。2
胞吞胞吐大分子和颗粒物质通过细胞膜包围形成囊泡,通过囊泡的形成和融合来完成运输过程,故被称作小泡运输(vesicular transport),又称批量运输(bulk transport),包括胞吞作用和胞吐作用。3
胞吞作用细胞质膜内陷将外来的大分子或颗粒物质包围形成小囊泡并转运到细胞内的过程称为胞吞作用(endocytosis)。根据胞吞作用对向的大小、状态机特异性,将其分为三大类:吞噬作用、胞饮作用及受体介导的胞吞作用。3
吞噬作用
吞噬作用主要是指某些特定的免疫相关细胞捕获较大的颗粒物质或大分子物质时,细胞膜凹陷,将颗粒包裹后,转运到细胞内,吞噬过程中细胞膜形成的小泡称为吞噬体(phago-some)或吞噬泡(phagocytic vesicle)。中性粒细胞、单核细胞及巨噬细胞等都具有吞噬功能,对颗粒物质的吞入是由质膜下微丝和结合蛋白的介导。这几种细胞广泛分布于动物体内,对入侵机体的病毒和细菌等微生物具有清除作用,对机体自身的代谢物如衰老死亡的细胞也具有清除作用。3
胞饮作用
当胞吞作用吞人的物质为液体时,称为胞饮作用(pinocytosis)。当胞外的溶质达到一定浓度时,可通过胞饮作用被摄人细胞内,这个过程中所形成的小囊泡的直径小于150 nm,小囊的形成受网格蛋白及结合素蛋白的介导。根据细胞外物质是否吸附在细胞表面,将胞饮作用分为两种类型。一种是液相内吞(fluid-phase endocytosis),这是一种非特异性的固有内吞作用,通过这种内吞作用,细胞把细胞外液及其中的可溶物吞入细胞内。另一种是吸附内吞(ab-sorption endocytosis),细胞外大分子或小颗粒物质先以某种方式吸附在细胞表面,因此具有一定的特异性。如小肠上皮细胞、血管内皮细胞等具有吞饮作用。3
受体介导的胞吞作用
通过与细胞膜表面特定受体结合,摄取细胞外蛋白、多糖或其他化合物的过程,称为受体介导的胞吞作用(receptor-mediated endocytosis)。因此,这种吞饮作用对摄入小泡的物质具有选择性,因此摄入的特异性强,效率更高。动物细胞对许多重要物质的吸收都是通过受体介导的胞吞作用来完成,包括多种蛋白质、生长因子、铁及维生素B12等。如动物细胞对卵黄蛋白的摄入过程,需要卵黄蛋白首先结合到细胞表面的受体,然后质膜凹陷将形成的卵黄蛋白及受体复合物包裹,形成小泡,随后小泡脱离细胞膜进入胞质,完成吞饮作用。此外,动物细胞通过受体介导的胞吞作用获取所需的大部分胆固醇。3
胞吐作用将细胞内的分泌泡或其他膜泡中的物质通过质膜运输到细胞外的过程称为胞吐作用( ex-ocytosis),是指大分子或颗粒状物质由细胞内排到细胞外的过程,分为连续性分泌和调节型分泌两种。连续性分泌( constitutive secretion)又称固有分泌,存在于所有真核细胞中,指分泌蛋白在内质网合成后,转运到高尔基复合体,并在此形成分泌泡,分泌泡与质膜融合,将分泌物排出细胞外。分泌的物质主要用于质膜的更新,包括膜脂、膜蛋白、胞外基质、营养及信号分子等。调节型分泌( regulated secretion)主要存在于特化的分泌细胞,分泌的蛋白合成之后被储存在分泌囊泡中,只有当细胞受到相应胞外刺激信号后,才启动胞吐过程。这种分泌方式只存在于分泌激素、酶、神经递质的细胞内,如血糖升高后,细胞会发出信号释放胰岛素,从而降低血糖。3
本词条内容贡献者为:
李丽霞 - 博士 - 西北农林科技大学
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国