

菊石目是头足纲、菊石亚纲的一目。壳多包卷,厚且多具显著饰纹,如横肋、纵脊、瘤、刺等。缝合线大都十分复杂,很少为二分叉的叶部或叶片状的鞍部。侏罗纪至白垩纪。除弛菊石目及叶菊石目外的所有侏罗白垩纪的菊石均属此目,包括属种极多。1
化石代表如香港菊石(Hongkongites)、束肋旋菊石(Virgatosphinctes)、白羊石(Ariecites)等。
发展历史菊石目是从约4亿年前的泥盆纪早期的册鹦鹉螺目进化而来的,在其后的3亿7千年间,在全世界的海洋里大量存在,直到白垩纪末突然消失。由于菊石进化很快、分布很广这一特点,这就使其在细分古生代晚期和中生代时期很有价值。作为一类,其特征是体管(与壳腔室相连接的管)的位置是在壳(腹)的外面。在古生代发现的菊石种的骨缝是单一的,而在中生代发现的则是复合的。由于它们已经绝种,我们对菊石的软体和生活习性知道得很少。保存的化石腔室中很少发现有咬颚、舌状、齿舌或墨液囊。
分类菊石目下已命名了四个亚目,分别为菊石亚目(下侏罗纪至上白垩纪)、勾菊石亚目(上侏罗纪至上白垩纪)、叶菊石亚目(下三叠纪至上白垩纪)及弛菊石亚目(下侏罗纪至上白垩纪)。不过《古无脊椎动物学论丛》却只包括了菊石亚目、弛菊石亚目及叶菊石亚目,并将之分类在菊石亚纲之下;而勾菊石亚目则被看为弛菊石亚目下的一个超科。2
另外有学者将这三个亚目分类在菊石目中。根据论丛所述,菊石亚目是于侏罗纪初期由叶菊石亚目及弛菊石亚目衍生而来。
形状尺寸 侏罗纪早及中期的菊石目很少会超越直径23厘米的大小。到了侏罗纪晚期及白垩纪早期,就开始出现了一些较大的形体,如英格兰南部的泰坦菊石直径就达53厘米,而白垩纪德国的Parapuzosia seppenradensis就更有达2米的直径。
侏罗纪早及中期的菊石目很少会超越直径23厘米的大小。到了侏罗纪晚期及白垩纪早期,就开始出现了一些较大的形体,如英格兰南部的泰坦菊石直径就达53厘米,而白垩纪德国的Parapuzosia seppenradensis就更有达2米的直径。
北美洲已知最大的菊石目就是Parapuzosia bradyi,直径长1.37米。另外,在英属哥伦比亚发现了直径达2.3米的未确认种,一旦获得确认,就是已知最大的菊石目。
结构形状基本结构 菊石目壳有间隔的部份称为闭锥。闭锥包含了逐渐变大的住室,住室之间有一道很薄的间隔隔开。菊石目动物只住在最后及最大的住室中。当它渐长时,会在开口处增加较新及较大的住室。连室细管由菊石目的身体,穿过间隔至所有空置的住室。
菊石目壳有间隔的部份称为闭锥。闭锥包含了逐渐变大的住室,住室之间有一道很薄的间隔隔开。菊石目动物只住在最后及最大的住室中。当它渐长时,会在开口处增加较新及较大的住室。连室细管由菊石目的身体,穿过间隔至所有空置的住室。
利用主导的高渗透运输作用,菊石目可以将这些住室的水份抽走,从而控制壳的浮力在水柱中上升下沉。菊石目与鹦鹉螺类主要不同之处是在于其连室细管在沿腹面穿过间隔的,而鹦鹉螺类的则是位于接近中央位置。3
两性异形现今鹦鹉螺壳的大小会与性别有关:雄性的壳较细小,而雌性的壳较大。这种两性异形可能也在菊石目中出现,亦正好解释了它们大小的变异。雌性需要较大的壳可能是为产卵之用。
侏罗纪早期欧洲的对菊石就是其中一个例子。菊石目壳的两性异形只是发现。以往这种大小的差异被认为是互为近亲的不同物种。
形状分类大部份菊石目壳都是呈扁平旋状,但也有一些是接近笔直的(如杆菊石)、像腹足纲壳的螺旋(如塔菊石及伯斯特菊石)、部份展开及卷曲而成熟时是笔直的(如澳洲菊石)。部份及完全展开的形态于白垩纪初期开始变得多样化,并被称为“异形菊石”。
异形菊石中外观最极端及异常的算是日本菊石,它们就像是一团不规则的螺旋,没有明显对称的卷曲。不过当近看时,就可以见到日本菊石壳上有U形的三维网络。
菊石目壳的表面分野很大。一些表面除了生长线外就很光滑,就像是现今的鹦鹉螺;一些则可以有螺旋脊及肋骨,甚至有棘伸出。这类的菊石目属白垩纪较后期出现。
口盖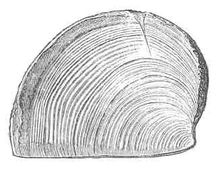 一些菊石目是连同一块角状板或一对方解石板一同被发现。以往认为这些板是用来开合其壳,就像是鳃盖一样。不过近却认为这其实是其颚部。这些板统称为口盖,单一块的称为单口盖,成对的则是双瓣颚。不论是单口盖或双瓣颚都很少能保存下来,而且都是与壳分离。
一些菊石目是连同一块角状板或一对方解石板一同被发现。以往认为这些板是用来开合其壳,就像是鳃盖一样。不过近却认为这其实是其颚部。这些板统称为口盖,单一块的称为单口盖,成对的则是双瓣颚。不论是单口盖或双瓣颚都很少能保存下来,而且都是与壳分离。
单口盖通常都是属于泥盆纪至白垩纪的菊石目,而双瓣颚则是属于中生代的菊石目。双瓣颚是对称的,大小及外观一样。口盖有多种形状,内外表面的纹刻也各有不同。
但是,由于它们很稀有,故仍不清楚哪一类口盖是属于哪一个物种,一些口盖甚至被立为独立的属。
软体部份虽然菊石目的化石数量很多,但其软体却十分稀少。
除了一些墨汁囊及可能的消化器官外,并没有发现任何软件部份。估计它们有很多触手来捕捉猎物。
生活习性菊石目及其近亲都已灭绝,保存下来的软体部份亦甚为稀有。大部份研究都是基于菊石目的壳及壳的模型。
大部份菊石目都可能生活在远古海洋的开放水域,而非海底。这是由于它们的化石往往都是在没有底栖动物的地层发现。很多菊石目相信都是游泳能手,如锐棱菊石,因它们的壳扁平、呈铁饼状及流线型,但也有一些可能是慢游或底栖的。
菊石目及其亲属可能会猎食鱼类、甲壳类及其他细小生物;它们的天敌包括水生爬行类,如沧龙科,在化石化的菊石目都有一些牙齿痕迹。它们可能会喷墨汁来逃避掠食,因为一些化石标本保留了这些墨汁。
菊石目的软体部份占有了壳末的最大节。较早而细小的节都有间隔,可以透过填充气体来维持浮力。故此较细小的部份会浮在较大的之上。
分布 由于菊石目可能是游泳或浮游的,它们往往都是栖息在海床以上的水域。当它们死亡后,就会下沉到海床及埋在沉积下,尸体会被细菌所腐化来维持平衡。
由于菊石目可能是游泳或浮游的,它们往往都是栖息在海床以上的水域。当它们死亡后,就会下沉到海床及埋在沉积下,尸体会被细菌所腐化来维持平衡。
很自然地,其化石周边就会出现矿物质的同心沉淀,称为结核,也是可以好好保存菊石目化石的原因。在黏土中发现的菊石目表面都会保存了珠母层的涂层,如英格兰的盔菊石。4
美国及加拿大的白垩纪皮埃尔泥板岩出产大量的菊石目化石,包括杆菊石、饼菊石、船菊石、装甲船菊石及Jeletzkytes,与及大量展开的形态。这些化石大部份都能保存原有的壳。
其他化石,如在马达加斯加及加拿大艾伯塔省发现的,都会有虹色。这些虹色菊石目在打磨后都有宝石的质素。相信这种虹色在活生生的菊石目并不存在,而是在其壳上多了额外的一层。
于中生代以后,菊石目化石就开始减少,直到新世代就完全消失。最后的生存线是于6500万年前的白垩纪-第三纪灭绝事件。于白垩纪晚期的生物集群灭绝后就没有菊石目生存,但却因个体发生学的分野而存留了一些鹦鹉螺类。若菊石目的灭绝是因火球撞击,周边的浮游生物会严重消失,造成菊石目繁殖的灾难。
国内发现1987年中国公布,继发现世界罕见的阿谢尔期菊石之后,又在广西南丹暮腰剖面的阿谢尔期菊石群下的麦蜓带中采获新的菊石。新发现的菊石与同在该剖面发现的阿谢尔期菊石、萨克马尔期菊石以及亚丁斯克斯菊石,构成界线清晰、年代连续的各类化石层。发现如此丰富的浮游生物化石和底栖生物化石,构成如此完整清晰的化石层序,在中国境内是绝无仅有的,国外也不多见。
这一重大地质成果,为世界石炭——二迭系界线的划分提供了宝贵的资料。
传说于中世纪的欧洲,菊石目化石被认为是石化了的蛇,故又被称为蛇石。相传这是由怀比特的希尔达(Hilda of Whitby)或圣帕特里克等圣人事迹的证据。商人会在菊石目的开口刻上蛇的面。
菊石目在尼泊尔卡格里达金河(Gandaki river)被称为“萨里格拉姆”,印度教相信它们是毗湿奴的显现。
本词条内容贡献者为:
胡芳碧 - 副教授 - 西南大学
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国