

你是否曾被某种声音打动过?是雨水滴落、鸟儿欢唱,还是悠扬琴声、琅琅书声?
能够聆听,是生物演化最美妙的事情之一。但是,敏锐的听觉不是神灵的恩赐,而是和身体其他机能一样,需要经历漫长的演化才能最终获得。
从外耳进入,穿过窄窄的耳道,穿过鼓膜,到达中耳区就能够看到三块迷你的听小骨——锤骨、砧骨及镫骨。我们的耳朵之所以能得到声音的青睐,就与它们有关。
一般认为,哺乳动物中耳经历了从下颌中耳,过渡型中耳,到典型哺乳动物中耳的三个演化阶段。然而,中耳内的骨骼是如何与下颌分离的,各个演化阶段发生于何时,首先发生于哪些哺乳类动物之中,为何会促使这样的分离?这一系列问题都让我们感到好奇。
11月28日,英国《自然》杂志在线发表了来自中科院古脊椎所王海冰、王元青以及美国自然历史博物馆孟津团队的研究成果。他们从一种新发现于辽宁凌源的早白垩世古老哺乳动物——盖氏热河俊兽(Jeholbaatar kielanae)的头颅里,获取到了早期哺乳动物中耳演化的新信息,为研究早期哺乳动物耳区演化提供了直接证据。

新的研究,讲述了一段发生在现生哺乳动物听力形成之前、关于最早获得典型哺乳动物中耳形态的演化故事。

盖氏热河俊兽生态复原图(图片来源:中科院古脊椎所许勇 绘)
原始哺乳动物的“耳朵”长啥样?
在现生的爬行类中,中耳区只有“耳柱骨”,我们人类的听小骨——砧骨、锤骨,也就是它们的方骨、关节骨,是连接爬行类动物齿骨与头骨之间的骨骼,并不发挥听力的作用。
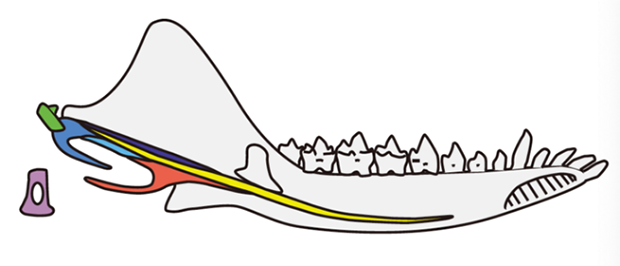
摩根齿兽的下颌中耳形态:耳区骨骼(齿骨后骨,postdentary bones)嵌入到下颌内后方(图片来源:Wang et al., 2019)
目前发现的早期哺乳型类动物(Mammaliaformes),比如摩根齿兽、柱齿兽类、巨颅兽,保持着比较原始下颌中耳状态(目前在柱齿兽类、巨颅兽中没有中耳骨骼保存,但从下颌骨内侧的形态,大致可以推断这些动物都具有下颌中耳)。
哺乳动物中耳演化的下一个阶段,**中耳形态发生了明显的变化,中耳骨骼变小、逐渐脱离下颌骨,并通过骨化的麦氏软骨与齿骨相连,这种形态被称为过渡型中耳。**在真三尖齿兽类中,就能窥见这一演化进程的秘密。
其他部位的骨骼是如何“转正”变成听小骨的?
时光回到2011年,同样在《自然》杂志,发表本篇论文的研究团队了报道了一种新的真三尖齿兽——胡氏辽尖齿兽。它的外鼓骨、锤骨、砧骨已经不再直接与齿骨接触,而是通过骨化的麦氏软骨与齿骨相连,也就是齿骨后骨开始转变为现生哺乳动物中耳的听小骨。

胡氏辽尖齿兽复原图(图片来源:ivpp)
胡氏辽尖齿兽发现于辽宁的白垩纪地层中,属于热河生物群中的一员。时隔几年之后,该团队又在辽宁凌源下白垩统九佛堂组地层中,发现了一种新的多瘤齿兽类——盖氏热河俊兽。它的属名取自热河生物群(Jehol Biota),种名是为了纪念波兰古生物学家索菲娅?盖兰-娅瓦洛夫斯卡Zofia Kielan-Jaworowska。
此次新发现的标本,与辽宁北票鲟化石保存在同一块岩板上。和庞大的鲟鱼相比,家鼠(体长约15厘米)大小的盖氏热河俊兽,蜷缩在角落里,显得弱小而孤寂。幸运的是,这个小小的身躯——盖氏热河俊兽的正型标本,却保存了完整的中耳形态,曾经属于头骨的方骨、下颌的关节骨,都跑到了它的中耳区,并且完全脱离齿骨,显示出“典型哺乳动物中耳”特征。

与北票鲟同眠的盖氏热河俊兽 (图片来源:论文作者提供)
多瘤齿兽,与恐龙同行
多瘤齿兽,顾名思义就是一种牙齿冠面布满数列瘤状的早期哺乳动物。多瘤齿兽类可谓是中生代哺乳动物中最大的类群,最早出现在中侏罗世,因为其生态方式与啮齿类动物相似,曾被称作“中生代的啮齿类”。
实际上,它们不止生活在中生代,而在白垩纪末幸运地逃脱了与非鸟类恐龙一起灭绝的厄运,以高度多样化的生存方式(包括体型的变异和牙齿的复杂化),一直幸存至始新世末,才退出生命演化的舞台。
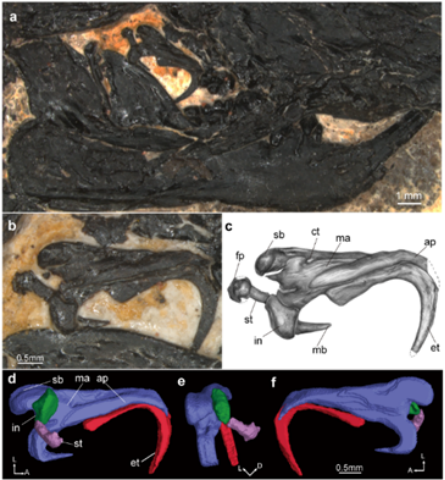
热河俊兽的左侧中耳形态(图片来源:Wang et al., 2019)
盖氏热河俊兽属于多瘤齿兽类,除了具有多瘤齿兽的基本特征之外,得益于化石的完整保存,以及高精度CL(Computed Laminography)扫描的运用,研究人员获取了热河俊兽所代表的多瘤齿兽类的中耳立体结构。其中,锤骨、砧骨的形态完整,基本保留了原始关节状态,二者呈叠覆型(背-腹型)的接触关系。

呈叠覆型(背-腹型)接触关系示意图
(黄色箭头示意接触方式,绿色代表砧骨,蓝色系代表锤骨)
(图片来源:Wang et al., 2019)
这种接触关系,与现生的单孔类以及已经灭绝的真贼兽类(如树贼兽)相似,而与现生兽类(有袋类和有胎盘类)和真三尖齿兽类(如胡氏辽尖齿兽)不同,后二者所呈现的是鞍型关节。实际上,研究人员发现,尽管中耳骨骼的形态变化很大,但锤骨-砧骨的关节方式一般都呈现出这两种结构。
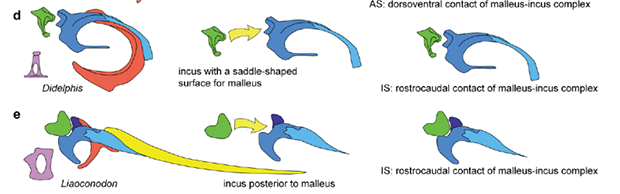
鞍型关节接触关系示意图,d负鼠;e辽尖齿兽
(黄色箭头示意接触方式,绿色代表砧骨,蓝色系代表锤骨)
(图片来源:Wang et al., 2019)
此外,本次研究还意外地发现,上隅骨从一块独立的骨骼逐渐变成与锤骨体愈合的状态,成为锤骨的后外侧部分。这为中耳骨骼的同源性讨论提供了重要证据。
结合详尽的形态观察与对比分析,研究人员最终确认了盖氏热河俊兽已经具有“典型哺乳动物中耳”这一事实。那么,接下来要解决的问题就是,这一演化过程,是如何发生的?
取食方式,领衔中耳演化进程
依托中国特有的燕辽、热河生物群中产出的哺乳动物化石,近几年来对异兽类(多瘤齿兽+贼兽类)的研究取得了一系列进展。
如今可知,至少在中/晚侏罗世(大约1.6亿年前),异兽类就已经演化出典型哺乳动物中耳。而在同一时期,甚至更晚的早白垩世(大约1.2亿年前),其他已知的所有哺乳动物类群都还保留了过渡型中耳。
那么,**为什么会演化出典型哺乳动物中耳呢?**学术界一直存在两种假说。
一种为“脑颅膨胀”假说,即哺乳动物生长过程中脑颅的增大导致中耳位置后移,最终脱离下颌。另一种为 “负向异速生长”,即在胚胎发育早期中耳骨骼形态相对于下颌较大,中耳骨化的时间更早,因此在胚胎发育后期,随着头骨、下颌的增大,中耳骨骼最终脱离下颌。
本论文提出了第三种假说:听小骨最终脱离下颌,可能与下颌的运动方式有关。也就是说,和它们的取食方式有关。
研究人员发现,在包括热河俊兽在内的异兽类,它们的齿骨-鳞骨颌关节方式独特,较为开放,能够支持下颌大幅度前后向的活动,与此同时,锤骨-砧骨所表现出的叠覆型的原始颌关节能够减少中耳骨骼在空间上的限制。
简而言之,热河俊兽的下颌骨能够做出比较大幅度的“前伸-回撤”动作,而为了给这种动作的运行幅度提供更大的空间、以便于它们取食,同时减少取食过程对听觉的影响,中耳骨骼脱离下颌而移入中耳区的时间,就被大大地缩短了。
所以,在“贪食”的选择压力之下,异兽类在至少1.6亿年前(早于其他所有哺乳动物类群),就已经为演化出“典型哺乳动物中耳”迈出了步伐。
不积跬步,无以至千里。虽然在我们人类的骨骼系统中,**听小骨是其中最小的骨骼,但是这一微小的结构让我们获得了更为敏锐的听力,以及对更大声阈范围的感应能力。**同样,最早演化出典型哺乳动物中耳结构的异兽类,凭借这一演化优势,为自己的夜间活动带来了更大的便利和安全保障。
想必,听力的提高,可能正是哺乳动物能够躲过中生代大灭绝、繁衍至今的关键原因之一吧。
参考文献:
-
Wang, H., Meng, J. & Wang, Y. Cretaceous fossil reveals a new pattern in mammalian middle ear evolution. Nature (2019) doi:10.1038/s41586-019-1792-0
-
Meng J, Wang Y, Li C. Transitional mammalian middle ear from a new Cretaceous Jehol eutriconodont[J]. Nature, 2011, 472(7342): 181.
-
Han G, Mao F, Bi S, et al. A Jurassic gliding euharamiyidan mammal with an ear of five auditory bones[J]. Nature, 2017, 551(7681): 451.
-
Benton M J. Vertebrate palaeontology[M]. John Wiley & Sons, 2014.
-
维基百科
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国