

奥陶纪是距今4.85-4.43亿年前古生代第二个时代,紧随寒武纪之后。奥陶纪是地史上大陆地区遭受广泛海侵的时代,是火山活动和地壳运动比较剧烈的时代,也是气候分异、冰川发育的时代。
奥陶纪时代,脊椎动物中的无颌类如甲胄鱼类等均已出现,但海洋中真正的主角还是海生无脊椎动物,海洋无脊椎动物在那个时期获得了极大发展,是海洋生物最主要的类群。有意义的是,在世界范围内的生物辐射事件再次发生,即所谓的**“奥陶纪生物大辐射”**。
在演化层次和分类等级上,这次大辐射虽逊于“寒武纪大爆发”,但其辐射规模远大于后者,是寒武纪的3倍。寒武纪生命大爆发主要体现在门纲级别的生物大辐射,而奥陶纪大辐射则在目、科、属级别的生物获得极大丰富和发展。
奥陶纪动物群主要由钙质壳腕足动物、三叶虫、半索动物、棘皮动物、苔藓动物、刺胞动物等构成。并且,奥陶纪增加了大量底栖游移、浮游生态分层和群落复杂性,食物链更趋复杂和完善。
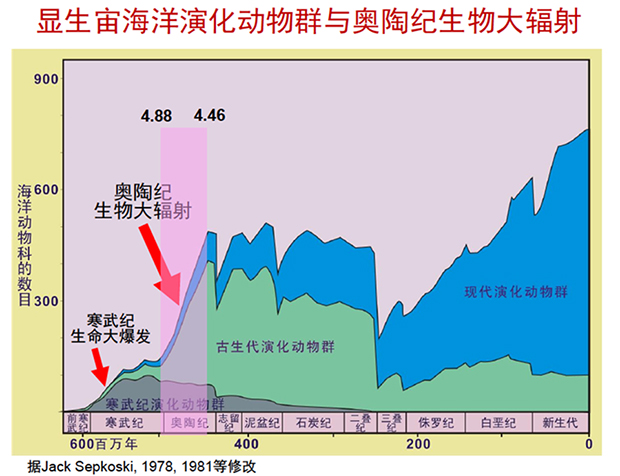
图1显生宙海洋演化动物群与奥陶纪生物大辐射(图片来源:据詹仁斌2017年对JACK SEPKOSKI, 1978,1981等修改)
各类无脊椎动物“明星”在奥陶纪竞相发展
奥陶纪海生无脊椎动物在浅海中空前繁盛,在海洋生态系中占据了主导地位,尤其以鹦鹉螺为代表的头足动物逐渐取代了奇虾,成为海洋中的新霸主,处于食物链的顶级位置。
鹦鹉螺动物最长可达4米,是海洋生物诞生以来“身体”最大的动物,是奥陶纪名副其实的海洋巨无霸。其头的两侧有眼,中央为口,口内有颚和齿舌,像鹦鹉的嘴,故称其为鹦鹉螺;口的四周有很多长的腕,腕上有吸盘,可捕食也可爬行。鹦鹉螺类始于寒武纪晚期,至奥陶纪最盛。

图2 奥陶纪巨无霸鹦鹉螺(图片来源:南京古生物博物馆)
笔石动物显得精致可爱。其形态十分奇特,有的呈树状,有的如展翅飞翔的大雁,还有的像张开的弓,但最奇异的还要数那些表面布满各种网眼、中间围成一个空腔的网兜状的细网笔石。
笔石与现今海洋中的杆壁虫十分相似,可能属于半索动物。笔石有两种生活方式,原始的笔石动物就像一棵树一样,固着在海底,然后向上生长;比较先进的笔石动物主要是营随波逐流的漂浮生活,依靠笔石虫体的触手摆动,滤食海水中悬浮的有机物。奥陶纪和志留纪是笔石最繁盛的时代。有时在一块标本中就能见到数以百计的笔石个体动物,足见在当时,笔石动物在海洋中是何等的丰富。

图3 笔石化石(图片来源:张元动提供)
三叶虫在寒武纪时就已经非常繁盛,奥陶纪时期进化出了新的类型。在海洋底层生活的三叶虫身体扁平,头部结构坚硬,便于挖掘;由于大量食肉类鹦鹉螺的出现,为了防御,有些三叶虫在胸、尾长出许多针刺;另外一些三叶虫,还进化出了非常巧妙的脊椎似的结构,用以抵抗刚出现不久的天敌——鱼类的捕食。
奥陶纪牙形动物极其多样,达到了历史发展的巅峰。那时的海洋乃是牙形动物的乐园,生活着大批的牙形动物。
奥陶纪腕足动物也极为丰富,属的总数是寒武纪的6倍。中国华南资料表明,腕足动物辐射始于早奥陶世晚期浅海细碎屑底质区,后在晚奥陶世初占领较深的海水环境。

图4 腕足动物化石(图片来源:詹仁斌提供)
这一时期海洋生态系统更趋多样化
奥陶纪的海洋生物类别繁多,与栖息地多样化密切相关。在台地环境,有深层掘穴的双壳类、浅层掘穴的多毛类、部分腕足类和三叶虫、低位固着底栖的腕足类、双壳类等、高位固着底栖的有柄类棘皮动物、苔藓虫、珊瑚、树形笔石、卧栖的软舌螺、底栖游移的三叶虫、双壳类、腹足类和介形类等,形成从海底之下到海底之上不同高度的生态分层。
此外,在陆棚水域、远洋水域和斜坡环境,还有游泳的头足类,以及浮游(或漂浮)的笔石动物、疑源类、几丁虫和放射虫等。因此,奥陶纪时期从海底向上到海洋表层水,从滨岸到深水斜坡地带,生态空间均被有效占领,每一种生态空间中都有相当丰富的不同门类的生物。
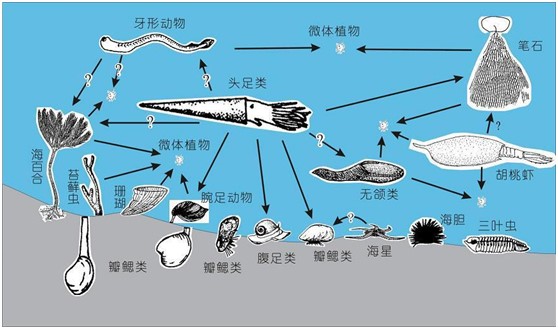
图5 奥陶纪海洋生态关系食物网(图片来源:张元动等,2009)
在奥陶纪各种生态类型的生物中,以底栖固着类型占优势地位。如大量爆发的腕足动物、棘皮动物、苔藓动物、珊瑚和海绵动物等,它们在海底之上的不同水层活动和取食。同时,内栖生物也开始增多,有些动物(如瓣鳃类)可以掘穴或钻孔到海底之下超过12 cm的深度。
在奥陶纪,游泳和浮游生物开始崛起,如大型游泳生物——头足类和浮游的笔石动物,它们演化迅速,在不同深度的海水生态空间中均占有重要地位。
食物链关系“重新洗牌”
随着奥陶纪时代的来临,生物界业已建立起来的食物链发生了很大的变化。位居食物链顶端的肉食动物不再是寒武纪时代称霸一时的奇虾,随着奇虾的没落,取而代之的是体型更大、游泳更快速和适应环境更强的肉食头足类,如直角石头足类,它们的后代就是大家十分熟悉的现代海洋中的章鱼、乌贼、鹦鹉螺,它们同样以超强的游泳和捕食能力占据海洋霸主的一席之地。
在摄食方式方面,奥陶纪海洋生物最大的特点是涌现出了大量的、多样化的滤食性生物。
它们以悬浮物或小型浮游生物为食,如部分瓣鳃类、喙壳类和磷质壳腕足动物在海底浅层滤食,多数瓣鳃类、单板类和腹足类在海底表面移动滤食,钙质壳腕足动物等用纤毛环在海底之上低位滤食,珊瑚和苔藓虫等用触手在海底之上的不同高度摄食,海百合和海林擒等由于长茎的存在得以用触手在海底之上高位摄食(25-100cm),笔石动物等则在开阔和远洋水域用纤毛触手滤食,从而形成海底之上不同高度的摄食生态分层。大量滤食性生物的爆发和摄食生态分层的扩展是奥陶纪生物大辐射的一个重要特征。
奥陶纪增加了一些肉食性动物,如远洋和开阔水域的头足类、肢口纲动物、无颌鱼类,以及海底游移的海星等,其中部分直接用各种捕食器官主动捕食。
此外,植食性生物也显著增加,除寒武纪已出现的单板类和介形虫外,奥陶纪增加了海胆、部分腹足类、介形虫和软甲纲节肢生物(如胡桃虾)等,以海底的微体植物为食。与此同时,以海底沉积软泥为食的生物继续繁衍,如三叶虫、部分腹足类和介形虫等。
总体而言,许多海底和远洋食肉类动物的出现和植食性动物类群的增加,使得奥陶纪的海洋生物在食物结构方面更加完善。
因此,奥陶纪海洋生物很可能已形成一个较为稳定、复杂的食物网或食物链。
根据对笔石胞管口饰的功能分析,专家推断,笔石可能是许多其他化石门类的食物,其中最可能的捕食生物包括头足类和节肢动物等。近年来在我国浙江和瑞典发现的笔石伤残标本,以及在美国奥陶纪和瑞典志留纪地层中发现的具疱状寄生痕迹的笔石标本,为笔石与其他生物的共生关系提供了新的证据。
在三叶虫的食性中,捕食是一种原始方式,主要局限于寒武纪早期的类型。奥陶纪的类型主要包括食颗粒物和滤食等,所以尽管奥陶纪的三叶虫可以在海底自由移动,但它们很可能不是捕食者,而更可能是被捕食动物。
奥陶纪的腕足动物多为海底固着,以滤食为生,也更可能成为被捕食对象,美国晚奥陶世地层中的腕足动物化石保存了它们被头足类捕食后留下的伤疤。浙西、赣东晚奥陶世腕足动物群中还保存了角环石寄生在活体腕足动物壳上的实例。
此外,头足类捕食腕足动物的证据也见于美国石炭纪地层记录,二者之间捕食与被捕食的关系可能早在奥陶纪就已建立起来。在瑞典,奥陶纪腹足类也有被捕食后造成的伤痕证据。
地形、地质和洋流变化是奥陶纪生物大发展的原因
奥陶纪是继寒武纪生命大爆发后又一次生物大发展的时代,尤其以无脊椎动物竞相发展为显著特点。那么,究竟是什么原因促使生物大发展的呢?
科学家近来的研究表明,早奥陶世随着温度升高,海面上升,扬子台地逐渐被淹没。紫红色、含氧量高的泥质灰岩沉积覆盖了整个台地,氧化的同时,将**原有的台地地形****“改造”**为较为平缓的斜坡,增加了不同的生态位,为相关的生物提供了生存活动的空间。
此外,由于区域地质构造运动及风化作用,一些来自陆地的岩石碎屑进入海中,增加了海底的养料,从而为不同种类生物的生存提供了适宜的环境条件和丰富的营养来源,导致了奥陶纪第一次生物大发展。
后来,伴随着晚奥陶世华南板块向北漂移,冈瓦纳大陆沿岸存在一个从南极到赤道的冷洋流,又称为“冷舌”。受其影响,冷水洋流给华南板块带来了丰富的营养物质,促进了海洋生物群的分异和繁盛,从而发育了凉水型动物群,记录它们的就是一套凉水型碳酸盐岩。因此,赤道“冷舌”是华南奥陶纪生物大辐射的背景机制之一。
总之,奥陶纪是海洋无脊椎动物时代,它开启了古生代生物大发展的大幕。
参考文献:
1、 詹仁斌,靳吉锁, 刘建波. 2013. 奥陶纪生物大辐射研究: 回顾与展望. 科学通报, 58(33), 3357–3371.
2、 戎嘉余, 黄冰. 2014. 生物大灭绝研究三十年. 中国科学 地球科学, 44, 1–28.
3、 张元动,詹仁斌,樊隽轩,成俊峰,刘晓. 2009. 奥陶纪生物大辐射研究的关键科学问题。中国科学 D辑:地球科学,39,129-143.
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国