

很长一段时间里,我们认为,生命遵守着它自己的一套法则,但当简单系统表现出“类生命”行为时,科学家开始为此争论——这些显而易见,又难以解释的现象背后是否都是热力学在作祟?

物理学与生物学之间有何不同呢?
当我们把一个高尔夫球和一颗炮弹同时从比萨斜塔上释放,根据物理学的法则,你可以非常精准地预测它们的轨迹。现在,我们用一只鸽子取代炮弹,重复刚才的实验(鸽子会飞走)。生物学系统不会违背物理学定律,当然,它们似乎也不具有预测的能力。相比之下,它们的目标直截了当——生存和繁衍。
我们可以认为,生物是有目的地引导它们的行为,或者用哲学家常用的话来说——目的论。

出于相同的原因,现在的物理学让我们可以从宇宙大爆炸后的十亿分之一秒来预测今天世界的模样。但是,没有人能想象,地球上的早期最原始的细胞是如何演变成人类。由此看来,物理学定律似乎并不能主宰生物的进化。
进化生物学家恩斯特·迈尔(Ernst Mayr)认为,目的论和生物学中的历史偶然性让生物学在众多科学中变得独一无二**[1]****。**这两项特性可能也都来源于生物学中的唯一指导法则——进化。
进化的内在虽然依赖于机会和随机性,但自然选择却赋予了它目的和意图的外表。动物们会被水吸引,并不是因为磁力的作用,而是出于它们的本能——求生意图。在这个过程中,腿是目的的体现者,并带着它们到达水源地。
迈尔声称,这些特性让生物学成为特例,即生物学本身就是一个法则。但最近,非平衡物理学中,复杂系统科学和信息理论的发展对此观点提出挑战。
一旦我们将生命看成可执行的计算代码(即在不可预测的环境中收集和储存信息),并考虑复制、适应、媒介、目的和意图,那么就可以理解,生物并不是由进化即兴产生,而是由于物理学法则产生的必然结果。
换句话说,似乎是有种物理学的东西在搞事情,并推动事物的演化。那么,一度被认为是定义生命系统特征的目的和意图,也许会通过热力学统计物理自然产生。

2016年11月,美国新墨西哥州圣菲研究所的一个研讨会[2]汇聚了一大批研究“复杂系统”科学的人,物理学家、数学家和计算科学家与进化分子生物学家就这些问题进行了会谈和争论——究竟生物学是否特殊?它的特殊之处在哪里?
毫无疑问,他们并没有达成共识。但却释放了一个很清楚的信号,如果在生物目的论和媒介的背后存在某种物理学机制,那么,它就会与近乎已成为基础物理学核心概念的信息有某种相关性。

无序和麦克斯韦妖
第一次把信息和意图考虑进热力学法则是在19世纪中期,那时候,统计力学的开山鼻祖——苏格兰科学家詹姆斯·克拉克·麦克斯韦(James Clerk Maxwell)正开辟统计力学的时代。麦克斯韦展示了如何引入信息和意图,并去做被热力学认为不可能成功的事情。
那时,麦克斯韦已经证明了气体属性,包括气压、体积和温度,是如何从无数随机和未知的分子热运动中,得出它们之间满足的可靠数学关系。换句话说,热力学是分子和原子在微观尺度统计力学下的产物,这门关于热运动的新科学,它连接着大规模物质的属性,比如气压和温度。
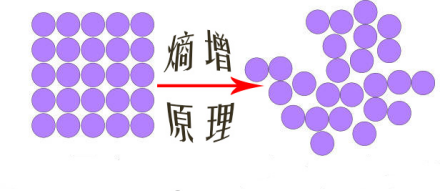
根据热力学规律,我们从宇宙能源中提取的、有用功的容量总是在减小的。随着有用的能量一直减小,热浓度总是趋向于变得平缓。在每个物理过程中,一些能量会通过分子间的随机运动,不可避免地成为无用热量而耗散掉。
这种随机性等同于一个很有意思的热力学量——熵(表征系统的无序性),也即热力学第二定律。最终,所有宇宙中的事物将会变成一种均匀、杂乱无序的状态——热力学平衡态,其中熵会达到最大值,到那时,一切有意义事物都不复存在。
难道我们真的注定是这种苦闷的命运?不,麦克斯韦不愿意去相信它,正如他后来提到的那样,1867年,他开始着手寻找热力学第二定律中的“漏洞”。他的目标从处理一盒无序并随机运动的分子团开始,接着他把运动快的分子从运动慢的分子中分离出来,从而减少这一过程中的熵。
想象一些可以看见盒子里每个独立分子的小生物,不像麦克斯韦的那么沮丧,后来的物理学家威廉·汤姆森(William Thomson)称之为妖。这个妖通过一个滑动隔板门把盒子分成两个部分,每当他看到一个具有特定能量的分子从盒子的右方接近滑板门时,它就会把门打开,并让分子通过;而当它看到一个慢一些、“较冷”的分子从盒子的左侧接近滑板门时,它也会把门打开,让分子通过。
最终,它会得到一个右侧部分是冷气体而左侧部分是热气体的盒子,这就是储热器工作的原型。
这种情况发生必须具备两个条件:第一,这个妖要比我们拥有更多的信息,因为它可以看到每个分子个体的运动状态,而不是统计平均值;第二,妖具有意图性——它可以有计划的把热分子和冷分子隔离开来。通过利用它有意图的学识,就能无视热力学定律。
人们花了一百年的时间去理解为什么麦克斯韦妖无法从事实上打败热力学第二定律,为什么麦克斯韦妖无法避免残忍的滑板门滑向死亡般普适的热平衡状态。
并且,有理由表明,热力学和信息运动过程中存在着更深的关联,也就是运算。美籍德裔物理学家罗尔夫·兰道尔认为[3],即使妖可以在无能量消耗的情况下,把信息和光滑(无摩擦)门的运动统一起来,最终总会有其他的东西为此买单。因为在它开始继续收集能量前,它不可能拥有每个分子运动无限制的记忆,它必须偶尔清除一下记忆,忘掉那些以前看到的,并开始新的记忆。而这种信息的擦写不可避免的代价就是需要消耗能量,因此,熵还是会增加。
所有违反热力学第二定律,依靠麦克斯韦妖灵活手工作业的努力,都被“兰道尔极限(信息擦除需要有限成本)”给消灭了。(更普适一点来讲,信息从一种形式转换成另一种形式是需要成本的。)。
**生物体似乎更像麦克斯韦妖。**虽然一个充满化学反应物质的烧杯,最终会消耗掉它的能量并进入到无聊、停滞的平衡态,但是从大约35亿年前生命的起源开始,生命系统就开始有组织地避免落入无生命性的平衡态。
他们有“目的”地从周围的非平衡状态环境中获取能量,即使是简单的微生物——细菌,也会有“目的”地朝着热源和营养物质的方向运动。埃尔温·薛定谔(Erwin Schrödinger)在他1944年出版的《生命是什么》中,曾这样表述过:生命体是依靠“负熵”为食。

薛定谔说:“通过俘获和储存信息,它们做到了。”其中,有些信息是编码在它们的基因里面,并且传递给下一代,包括那些获取负熵的指令。
作为一个训练有素的科学家,薛定谔并不知道,信息是如何存储和编码的,但正是这个被他称之为“非周期性晶体”的直觉,却激励[4]了弗朗西斯·克里克(Francis Crick)和詹姆斯·沃森(James Watson)。并于1953年,他们发现了遗传信息是如何编码在DNA分子结构上的。

从那以后,基因组至少可以看作生物体祖先(可以追溯到很久以前)在地球上生存的一种记录。
根据大卫·沃伯特(David Wolpert)(数学和物理学家)和他的同事艾米蒂·科林斯基(Artemy Kolchinsky)在圣菲研所研讨会上的结果,有个关键点被提出来——**具有高适应性的生物与它所生存的环境相关联。**如果一个细菌是因为左边或者右边有食物而朝该方向游动,那么它就能比那些随意游动、全靠运气寻找食物的细菌具有更好的适应性和繁殖能力。这种存在于生物体和它所处的环境之间的关联性,表明它们之间的信息是共享的。
沃伯特和科林斯基认为,正是这种信息帮助了生命体保持非平衡状态,因为,就像麦克斯韦妖一样,它能从周围环境的波动中提取信息并确定它的行为。如果没有掌握这些信息,生物体就会慢慢地恢复到平衡状态——死亡。
以这种方式来看,生命可以被视为一种最大化储存和使用有用信息的运算**[5]**,而且生命也很好的证明了这一点。兰道尔的结论给麦克斯韦妖的难题设定了一个绝对下限——有限记忆的运算需要一定的能量,也就是说遗忘是耗能的。
当今最好的计算机所浪费的能量,要远远高于传统的能量消费和耗散,超了一百万倍还不止!但沃伯特表示:“我们可以对一个细胞完成总计算所消耗的能量和相应的热力学效率进行非常保守的估计,它仅仅是兰道尔极限值的十倍左右”。

无生命的进化论
他还说:“自然选择一直非常关注运算所需的最小化热力学成本。它会竭尽所能地去减少细胞必须执行的运算总量。”换句话说,生物(可能除了人类以外)似乎并不会在生存问题上过度关心。
他还表示,这种关于贯穿生物整个生命的能量损耗和运算的问题,一直都被目前的生物学所忽略。
生物体可以被看成一种实体,原因是它们可以通过使用信息来获取能量和逃避平衡态,从而适应周围环境。当然,这听起来有点绕口。但值得注意的是,这里并没有提到基因和进化论,而像迈尔等科学家却假定它们是生物的意图和目的所依赖的。
这样的图像能带领我们走多远?被自然选择过的基因无疑是生物学的核心。但自然选择过的进化论本身可能是存在于纯物理世界当中的、一个更具功能性和明显目的性的特例吗?由此看来,这还只是个开始。
适应性一度被认为是达尔文进化论的特点。但是麻省理工学院的物理学教授杰里米·英格兰[6](Jeremy England)对此表示质疑,他认为,对环境的适应性也可以发生在复杂的无生命体系里。
相比普遍认为生物体所有的特点都是为了生存的达尔文主义,此处的适应性有了更加丰富的内涵。达尔文观点中的一个难题就是,除非进行追溯,否则没有办法定义什么是具有高适应性的生物体。
那些“最具适应性”的生物体表现出更好的生存能力和繁殖能力,但是你并不能预测什么样的适应性是有意义的。比如,鲸鱼和浮游生物都是具有很好适应性的海洋生物,但是在具体的适应方式上,彼此之间却没有什么明显的联系。
英格兰对于“适应性”的定义更接近薛定谔,甚至是麦克斯韦,即一个具有良好适应性的个体可以从未知、扰动的环境中有效地吸收能量。就好比有个人能站在左右摇摆的船上,而其他人却都跌倒了,因为她更能适应甲板的晃动。
通过在非平衡条件下运用概念模型和统计力学的方法,英格兰和他的团队认为[7],这些具有良好适应性的系统应该属于那些能从环境中吸收和耗散能量的体系,并在此过程中产生熵。
他还表示:“热扰动的物质经常会自发地变成那种更容易从随时间演化的环境中获取功的形状。”即复杂系统倾向于以惊人的易用性融入到这些高适应性的状态中。
这个过程中,没有涉及到对周围环境的逐渐适应(而对周围环境的逐渐适应却贯穿了整个达尔文机制——繁衍、变异和性状遗传),没有一点关于繁衍的事情!英格兰还说道:“让我感到激动的是——这意味着,当给予我们一定物理数量的可观测原始适应性结构时,我们发现它们并不需要生理意义上的父辈结构。你可以用热动力学去解释进化论中的适应性,甚至可以更特殊一点,不需要自我复制那一套,达尔文的逻辑法则也不再成立”。(只要问题中的系统足够复杂、多变和敏感就可以应对它所处环境中的扰动)
但是,**物理学和达尔文的适应性并没有冲突。**事实上,后者可以看作前者的一种特例。如果复制存在的话,自然选择就成了系统从环境中获取功(即薛定谔的负熵)的途径。自我复制实际上是一种可以使复杂系统更加稳定的机制。所以不要奇怪,这就是生物学的用途!
但是在非生命的的世界中,自我复制却不会经常发生,那些具有良好适应性的耗散结构倾向于有高度的组织性,比如沙纹和沙丘,它们都是风沙的杰作的结晶。从这种方式来看,达尔文的进化论可以看作一种更普适物理规律的实例,并且这种规律还掌控着非平衡系统。

预言机
这种适应扰动环境的复杂结构图像,同时也允许我们去推断这些结构是如何储存信息的。简而言之,只要是这种结构,不论是有生命的还是无生命的,都会被迫使用可用的能量,并且它们有可能成为“预言机”。

生物学体系会改变它们的状态来回应一些来自环境的驱动信号,这几乎是生命特征的一种定义。只要有事情发生,你就会做出反应。植物会向着阳光生长,也会产生毒素去回应病原体。这些环境信号是典型的不可预测信号,但是,生命体系会依据经验,从它们的环境中储存和使用这些信息来引导未来的行为。(在这种图像中,基因只是基本的、普通意义上的关键点)
尽管预测性无法选择,但根据夏威夷大学的教授苏珊·斯蒂尔(Susanne Still)和加利福利亚州前劳伦斯伯克利国家实验室的研究员加文·克鲁克斯(Gavin Crooks),以及它们同事的工作,对未来的预测似乎对处于随机性和扰动性环境中的任何高效能量体系是必不可少的[8]。
斯蒂尔和同事发现,存储过去信息所产生的能量损耗对未来的预测没有价值。为了使有效性最大化,系统不得不进行选择。如果它盲目地记忆所有发生过的事情,就会导致大量能量损耗掉。
另一方面,如果它一点也不储存它所在环境中的信息,它就会不断地去应对意外。斯蒂尔的合作者,大卫·西瓦克(David Sivak)(现任职于不列颠哥伦比亚省本拿比的西蒙弗雷泽大学)说道:“一台最佳的热力学机器必须让它的情怀达到最小化——关于过去的无用信息——来平衡记忆和预测性。”简单的说,它必须变得擅长获取有用的信息,也就是那些有可能对未来生存起作用的信息。
你所期望的自然选择青睐那些能够有效利用能量的生物。但是一些个别的分子生物学器件,比如我们细胞中的泵和电机,在某些重要情形下,它们是通过对过去的学习来预测未来。斯蒂尔还说到,为了获得它们惊人的效率,这些器件必须抹去它们到目前为止对世界表现形式的简单印象,从而使它们能够预测将来的事情。

热力学定义的死亡
在缺少进化和自我复制的情况下,即使生命体系中的基本信息处理性质已经被提出来了,通过非平衡动力学,你如果要想象更复杂的特性,如工具的使用,交流和社会合作,你必须把进化考虑进来。
算了,还是别考虑它了。这些行为通常被认为是高度发达和进化的生物的优势所在,包括灵长类和鸟类,它们可以由相互作用粒子构成的简单系统进行模拟。
这项技术的关键在于,系统需要约束条件的引导,即在给定的时间里,它会表现出使熵值的最大化(在这种情况下,可能需要对粒子所处的路径进行不同的定义)。
熵的最大化一度被认为[9]是非平衡系统的特性。但是在这一模型下,系统遵守着在固定的时间窗口内,让它的熵最大化的规则,并会延伸至未来。换句话说,它具有前瞻性。
事实上,这个模型遍历了粒子可能存在的所有路径,并迫使它们去适应产生出最大熵的那条。粗略讲,它倾向于使粒子以后运动具有最多可选路径的那条。
你也许会说,粒子系统经历着一种保障它今后行动自由性的需求,并且这种需求引导着它每时每刻的行为。研究并发展了这一模型的学者,亚历山大·维斯纳·格罗斯(Alexzandra Wissner Gross)(哈弗大学)和卡梅伦·弗莱尔(Cameron Freer)(麻省理工学院的数学家)把这种需求称为**“熵的因果力”**[10]。在使用计算机模拟特别设置过的圆盘状粒子到处移动时,这股力量会产生具有某种智能性的结果。
比如有种情形,一个大的圆盘可以从细管中通过“利用”小盘去衍生出新的小盘,这个过程好比工具的使用,而新盘的释放又可以提高系统的熵值。另外一种情形,为了能与更大的圆盘产生相互作用,两个分别处于隔离区域的圆盘会同时去摧毁大盘,这就是社会合作的体现。
当然,这些简单的相互作用方式对它们瞥见未来是有益处的。但从普适法则来看,生命却并非如此。那么,这种作用是如何与生物学相关联的呢?
虽然维斯纳·格罗斯表示,他已经开始致力于为“熵的因果力”确立一种实际的、合乎生物学规律的机制,但并没有弄清楚。与此同时,他还认为这种方法可能有实际的应用价值——给人工智能提供途径。他预言,相比直接从运算或者预测技术出发,先观察现象,后研究其背后的物理规律和约束条件会更快一些。换句话说就是,先找到你所想要的体系,然后再去解决它是怎样工作的。
**衰老也一直被认为是一种由进化决定的形状。**生物体要在有限的寿命里去创造机会繁衍后代,围着父辈转圈的子代不会待太长时间,它们就要去谋生和争夺资源。这似乎本该就是故事的一部分,但是希尔德加·梅尔·奥尔特曼(Hildegard Meyer Ortmanns)(德国不来梅雅可比大学物理学家)却不赞同,他认为,衰老的根本是由热力学信息控制的物理学过程,而不是生物意义上的。

视频里,David Kaplan 解释了熵增法则是如何驱动随机性进入到稳定、有序的生命结构
当然,这绝不是把东西用完了那么简单。梅尔·奥尔特曼说:“我们制造的大部分软物质材料会在它开始衰老前就推陈出新”。但是,这种更新的过程并不完美。热力学信息的复制要求必须在精确度和能量二者间折衷[11]。一个生命体的能量供应是有限的,所以错误一定会随着时间积累下来。此时,生命体不得不耗费大量的能量去修复这些错误。更新的过程最终会因为复制产生了太多的缺陷而无法正常运转,接着,死亡便降临了。
经验似乎证明了这一点。众所周知,培养的人类细胞似乎最多只能复制40到60次,接着就会停止复制,走向衰亡。(这被称为海佛烈克极限,Haylfick limit[12])最近对人类寿命的观察表明,为什么人类百岁之后不会存活太久[13]?这里面可能有些本质的东西。
对于处在扰动的非平衡环境中,系统表现出明显高效率的能量需求、组织需求和预测性,这是有推论的。我们自己就是这样一个体系,就好比我们的祖先都回到了最原始的细胞,非平衡热力学似乎要告诉我们,在这种情况下究竟什么才是重要的。
换句话说,行星上生命的出现,比如早期的地球,充满了像阳光和火山活动类型的能源,它们持续破坏平衡。开始时,似乎没有什么极其罕见的事情发生,正如很多科学家所假定的那样,但实际上它就是会发生。
2006年,圣菲研究所的艾克·史密斯(Eric Smith)和后来的哈罗德·莫罗维茨(Harold Morawitz)认为[14],非平衡热力学系统更可能是在有着益生元(被认为是生命产生所需的物质)的地球上催生出组织和复杂系统,并使地球远离平衡态,而不是把原始的化学材料简单的放入“温暖的小池塘”(查尔斯·达尔文是这么称呼的)里,用小火炖出来的。

在提出这一争论的前十年里,研究者进行了细节补充并深入分析。那些被恩斯特·迈尔认为是生物学必不可少的属性——目的和意图,可能是热力学统计物理所产生的必然结果。而这些一般属性可能会反过来,自发产生出某些东西,比如生命。
与此同时,天文学家告诉我们,在我们所处的星系里,估计有数以十亿计[15]的行星绕着其他星球运转着。它们有许多处于非平衡态,至少有些就像地球一样,并且,同样的规则也在那里发生着。
文章出处:https://www.quantamagazine.org/20170126-information-theory-and-the-foundation-of-life/
参考文献
[1] Mayr E. What makes biology unique?: considerations on the autonomy of a scientific discipline[M]. Cambridge University Press, 2007.
[2] David Wolpert, Michael Lachmann, David Krakauer, Manfred Laubichler and P S. Statistical Physics,Information Processing and Biology[R]. New Mexico: .
[3] Landauer R. Irreversibility and heat generation in the computing process[J]. IBM journal of research and development, IBM, 1961, 5(3): 183–191.
[4] Jerry Coyne. Crick writes to Schrödinger, 1953[EB/OL]. /2017-01-26. https://whyevolutionistrue.wordpress.com/2015/01/05/crick-writes-to-schrodinger-1953/.
[5] Rovelli C. Meaning= Information+ Evolution[J]. arXiv preprint arXiv:1611.02420, 2016.
[6] Wolchover N. A New Physics Theory of Life[EB/OL]. . https://www.quantamagazine.org/20140122-a-new-physics-theory-of-life/.
[7] Perunov N, Marsland R A, England J L. Statistical physics of adaptation[J]. Physical Review X, APS, 2016, 6(2): 21036.
[8] Still S, Sivak D A, Bell A J等. Thermodynamics of prediction[J]. Physical review letters, APS, 2012, 109(12): 120604.
[9] Jaynes E T. Information theory and statistical mechanics[J]. Physical review, APS, 1957, 106(4): 620.
[10] Wissner-Gross A D, Freer C E. Causal entropic forces[J]. Physical review letters, APS, 2013, 110(16): 168702.
[11] Barato A C, Seifert U. Thermodynamic uncertainty relation for biomolecular processes[J]. Physical review letters, APS, 2015, 114(15): 158101.
[12] Shay J W, Wright W E. Hayflick, his limit, and cellular ageing[J]. Nature reviews Molecular cell biology, Nature Publishing Group, 2000, 1(1): 72–76.
[13] Dong X, Milholland B, Vijg J. Evidence for a limit to human lifespan[J]. Nature, Nature Research, 2016, 538(7624): 257–259.
[14] Morowitz H, Smith E. Energy flow and the organization of life[J]. Complexity, Wiley Online Library, 2007, 13(1): 51–59.
[15] NASA. Billions and Billions of Planets[EB/OL]. /2013-01-03. https://www.nasa.gov/mission_pages/kepler/news/kepler20130103.html.
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国