

概述
肌醇磷脂(phosphatidylinositol,PI)又称磷脂酰肌醇,自大豆中提取,由磷脂酸和肌醇组成,是细胞内的信使物质,广泛存在于细胞膜与内质网上,代谢活跃,具有重要的生理功能。在肝脏及心肌中多是一磷酸肌醇磷脂,而在脑中多为二、三磷酸肌醇磷脂。具有极性头和非极性尾两性酯类,是细胞膜组分之一。
肌醇磷脂代谢 1953年Hokin等发现乙酰胆碱作用于胰腺细胞,在有32P_ATP存在下,32P很快与膜磷脂结合,特别是进入肌醇磷脂。后来他发现在神经细胞及脑组织中有肌醇磷脂酶C,它特异水解PI,产生DG及IP;在微粒体及线粒体中,在ATP 存在下,DG可很快合成磷脂酸( PA ),PA在ATP存在下,经CTP磷脂酸胞苷酸转移酶催化下可形成CDP 一DG。HoKin在1959年提出了Pl合成的循环设想。1
1953年Hokin等发现乙酰胆碱作用于胰腺细胞,在有32P_ATP存在下,32P很快与膜磷脂结合,特别是进入肌醇磷脂。后来他发现在神经细胞及脑组织中有肌醇磷脂酶C,它特异水解PI,产生DG及IP;在微粒体及线粒体中,在ATP 存在下,DG可很快合成磷脂酸( PA ),PA在ATP存在下,经CTP磷脂酸胞苷酸转移酶催化下可形成CDP 一DG。HoKin在1959年提出了Pl合成的循环设想。1
1969 年Thompson用脑观察Pl代谢,发现不仅 l参与反应,PIP及PIP2也与Pl同时改变。他分析了PIP及PIP2的脂肪酸,发现甘油的第一个碳上连结的是C18硬脂酸,第二个碳结合的是花生四烯酸,故认为PIP、PIP2都是从PI 磷酸化而来,分别由PI,PIP激酶的催化。不同组织细胞的PI和PIP激酶在亚细胞组分中分布不同。如视网膜,肿瘤细胞,PI激酶主要与质膜结合,PIP激酶主要在胞浆上清液;大鼠肝细胞的PI激酶在高尔基氏体膜和核膜,PIP激酶与质膜结合。细胞内不仅有使Pl磷酸化的激酶,也有使PIP2水解的磷酸单脂酶,变成PIP及Pl。所以细胞内依赖磷酸激酶及磷酸单脂酶维持着质膜由Pl,PIP及PIP2之间的动态平衡。1
PIP2经肌醇磷脂酶C水解后,可生成DG及IP3,DG可再合成PI,lP3经磷酸酶作用,依次从5、4、1位将磷酸水解下来,成Ins1,4P2,Ins1 P及Ins,IP3也可继续磷酸化,由激酶催化形成lP4,IP5及IP6,这些化合物的功能还有待研究。
作用肌醇磷脂信号系统以肌醇磷脂代谢循环为基础, 由多种磷酸磷脂酰肌醇分子和多磷酸肌醇分子及催化代谢的磷脂酶、激酶组成。该信号系统参与调节动、植物细胞生长发育及应答环境刺激等多种生理过程。2
肌醇磷脂对高脂血大鼠血脂水平有降低作用,同时具有抑制脂质过氧化的作用。3
植物体内肌醇磷脂代谢与渗透胁迫信号转导:磷酸肌醇代谢在生物感受胞外刺激及信号转导中起着重要的作用。在此,植物体内的肌醇磷脂代谢途径,PI-PLC/IP3途径和磷酸肌醇激酶与渗透胁迫信号转导之间的联系,以及肌醇磷脂代谢参与的渗透胁迫信号转导与ABA之间的关系。4
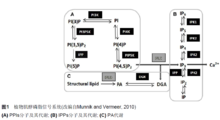 肌醇磷脂依赖性磷脂酶C(phosphatidylinositol-specific phospholipaseC, PLC)是肌醇磷脂信号系统中的关键酶之一。细胞受到外界刺激时,PLC被受体激活,水解膜组分PI(4,5)P2(phosphatidylinositol4,5-bisphosphate)产生双信使分子IP3(inositol 1,4,5-trisphosphate) 和DAG(diacylglycerol),调节细胞中Ca2+的浓度和PKC(protein kinase C)的活性。该信号途径即经典的双信使途径。多项研究表明,肌醇磷脂信号系统的多种组分参与了花粉发育过程中的液泡变化及花粉管生长中的囊泡运输过程。这些组分包括PI(3)P(phosphatidylinositol 3-phosphate)及催化其产生的激酶PI3K、PI(4,5)P2及催化其形成的激酶PIP5K、PLC/IP3/Ca2+、PA(phosphatidicacid)和PLD(phospholipase D)以及IPK (inositolpolyphosphate kinase)和IPP(inositol polyphosphatephosphatase)等。5
肌醇磷脂依赖性磷脂酶C(phosphatidylinositol-specific phospholipaseC, PLC)是肌醇磷脂信号系统中的关键酶之一。细胞受到外界刺激时,PLC被受体激活,水解膜组分PI(4,5)P2(phosphatidylinositol4,5-bisphosphate)产生双信使分子IP3(inositol 1,4,5-trisphosphate) 和DAG(diacylglycerol),调节细胞中Ca2+的浓度和PKC(protein kinase C)的活性。该信号途径即经典的双信使途径。多项研究表明,肌醇磷脂信号系统的多种组分参与了花粉发育过程中的液泡变化及花粉管生长中的囊泡运输过程。这些组分包括PI(3)P(phosphatidylinositol 3-phosphate)及催化其产生的激酶PI3K、PI(4,5)P2及催化其形成的激酶PIP5K、PLC/IP3/Ca2+、PA(phosphatidicacid)和PLD(phospholipase D)以及IPK (inositolpolyphosphate kinase)和IPP(inositol polyphosphatephosphatase)等。5
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国