

介绍
人类发现的第一个神经营养因子——神经生长因子( Nerve Growth Factor, NGF )首先是由意大利神经科学家 Rita Levi-Montalcini 和美国生物化学家 Stanley Cohen 于 1956 年分离成功; Cohen 还意外发现了另一种能促进表皮细胞生长、增殖和分化的生长因子,因而将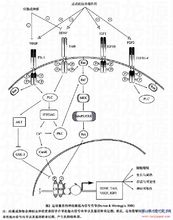 该因子命名为表皮生长因子( Epidermal Growth Factor, EGF )。为此, Levi-Montalcini 和 Cohen 于 1986 年共同获得了诺贝尔生理学或医学奖。
该因子命名为表皮生长因子( Epidermal Growth Factor, EGF )。为此, Levi-Montalcini 和 Cohen 于 1986 年共同获得了诺贝尔生理学或医学奖。
神经生长因子的发现过程可追溯到 20 世纪 20~30 年代。美国神经科学家 Samuel Randall Detwiler 和美国胚胎学家 Viktor Hamburger 那时就已发现神经元所支配的靶细胞在神经元存活中具有重要作用。他们观察到,两栖动物胚胎背根神经节中的感觉神经元数量可因移植外加的肢芽( limb bud, 指胚体外侧壁在将来发生四肢的区域向外膨出而形成的隆起)到靶细胞中而增加。相反,神经元数量可因去除正常的靶细胞而明显减少。他们认为,由靶细胞引起的神经元数量改变是靶细胞影响成神经细胞增生与分化的反映。但在 40 年代,这种解释受到意大利年轻的神经科学工作者 Rita Levi-Montalcini 的质疑,因为在她的实验中,她惊奇地发现在胚胎的发育过程中神经元死亡是完全正常的事。
发现过程1947 年秋, Levi-Montalcini 接受 Viktor Hamburger 教授的邀请前往美国参加他的工作,并重复她自己许多年前在鸡胚上所做的实验,这是 Levi-Montalcini 一生中的重要转折点,后来她在自传中如是写道。 在关键的实验中,她和 Viktor Hamburger 证实了与保留完整的肢芽的鸡胚相比,去除鸡胚的肢芽将导致更多的感觉神经元死亡。
在中枢神经系统的发育过程中,神经细胞也发生正常死亡。 Hamburger 后来证实在胚胎发育过程中在鸡脊髓外侧运动柱中生成的运动神经元约有一半是注定要死亡(凋亡)的。而且,在类似于对感觉神经节的那些实验中, Hamburger 发现运动神经元的死亡数量可因去除靶细胞而增加或因外加肢芽而减少。所以肌肉靶的大小对脊髓运动神经元的存活是极为重要的。神经元过度生成过程后的死亡几乎发生在中枢和周围神经系统的任何部位。
Hamburger 和 Levi-Montalcini 的上述发现促使他们提出了神经营养因子( neurotrophic factor )的假设,这个假设认为发育中的神经元所支配的靶细胞可产生和分泌一种数量有限的但又是必需的营养性物质或营养因子,而这种营养性物质或营养因子可在神经末梢被摄取,那些不能获得适量神经营养因子的神经元将会死亡(凋亡)。根据这一假设,最终 Levi-Montalcini 和她在 Hamburger 实验室的同事生物化学家 Stanley Cohen 一起成功分离了神经生长因子( nerve growth factor, NGF )。在这个过程中,有两个重要的事件值得一提。
第一个事件是在 Hamburger 和 Levi-Montalcini 证实靶组织具有调节神经元的存活数量后不久, Hamburger 以前的一个学生 Elmer Bueker 通过实验证明了不同的肿瘤组织移植至小鼠能替代外周的靶细胞支持感觉神经元的存活。 Bueker 发现小鼠肉瘤组织可使感觉神经纤维广泛长入肿瘤组织,他还观察到脊髓中植入肿瘤的一侧背根神经节明显大于另一侧相对应的神经节。这些成果被 Levi-Montalcini 和 Hamburger 进一步拓展,他们注意到在肉瘤植入近旁的交感神经节明显增大,并进一步证实肉瘤细胞的作用是由一种可扩散的因子所引起的。 Levi-Montalcini 研制了可定量分析肿瘤组织对离体的感觉神经节和交感神经节存活和轴突生长的方法。 1956 年,她和生物化学家 Stanley Cohen 一起提纯了这个可扩散的分子,并将这个分子命名为神经生长因子。
第二个事件是在一个关键的生物化学实验中, Cohen 和 Levi-Montalcini 为了排除 DNA 或 RNA 对神经产生营养作用的可能性。他们碰巧使用了一种粗制的蛇毒标本作为具有活性的磷酸二酯酶来降解存在于不完全纯化的 NGF 标本中的各种核酸,结果他们发现蛇毒本身具有比 NGF 更为显著的促轴突快速生长效应。然后, Cohen 研究了雄性小鼠的颌下腺,这是在哺乳动物相当于蛇毒腺的部分,结果令人兴奋地发现它富含 NGF ,这一结果启示了可供 NGF 纯化和蛋白测序的丰富来源。随后的工作证明了 NGF 是由 2 个 a 、 2 个 b 和 2 个 g 亚单位组成的蛋白质,其分子量约为 130000 。 a 亚单位具有胰蛋白酶样活性, b 亚单位的结构与胰岛素相似,而 g 亚单位则为丝氨酸蛋白酶。 b 亚单位是 NGF 促进神经生长的主要活性部分 ,是一种含 118 个氨基酸残基的序列 。
Rita Levi-Montalcini 于 1909 年 4 月 22 日 出生于意大利都灵市的一个 犹太人家庭。她的父亲是个电机工程师和有天赋的数学家,母亲是个有才华的画家。 1936 年, Levi Montalcini 毕业于都灵医学院,但由于当时墨索里尼法西斯独裁政权对犹太人的歧视政策,她的学术研究生涯不幸被暂时中断。她先在都灵自己家里坚持科学研究工作,她把她的实验室建在自己的卧室里。 1941 年因都灵遭受英美空军的空袭迫使她放弃都灵,搬迁到乡村农舍继续她的实验。 1943 年秋,德国军队入侵意大利,迫使她 和家人一起 逃往佛罗伦斯, 在那里他们隐蔽地居住到战争结束。 1945 年 5 月。 她和家人才回到都灵,并恢复了她在大学的科学研究工作。 1947 年秋接受 Hamburger 教授的邀请到美国圣路易斯继续她的研究,并在那里取得了举世瞩目的成就。她原本打算在圣路易斯逗留 10~12 个月,但由于出色的研究成果使她大大延迟了回意大利的时间。 1956 年,她被任为副教授, 1958 年起任正教授,直到 1977 年退休。 1962 年,她在罗马建立了一个研究所,此后就在圣路易斯和罗马两个城市之间奔波。 1969~1978 年间,她还担任意大利国家研究委员会细胞生物学研究所的主管, 1979 年退休后,她仍被聘为该研究所的客座教授。
分类和作用NGF 的发现是研究生长因子和激发寻找其他神经营养因子的里程碑。现已知道, NGF 仅仅是一系列具有促进神经元存活的分泌因子之一。研究最多的一类营养因子是神经营养因子(neurotrophins)。四种主要的神经营养因子已从哺乳动物中分离出来,它们是: NGF 、脑源神经营养因子(brain derived neurotrophic factor, BDNF),神经营养因子3(NT-3)和神经营养因子4/5(NT-4/5)。此外,可能还有神经营养因子6(NT-6)。1
NGF 广泛存在于人和多种动物体内。若将 NGF 的抗血清注入新生动物,可使几乎所有的交感神经节受损;而将抗血清注入母鼠则可导致胎鼠感觉神经系统缺失。可见 NGF 对交感神经元和感觉神经元的生长和存活是必需的。实验还表明, NGF 能提高基底前脑和纹状体胆碱能神经元的 cAMP 水平,增高胆碱乙酰移位酶的活性,并对这些神经元的生长和存活起重要作用。各种 NT 在保护神经元和阻止细胞凋亡中有不同的作用,但也有一些交叉和重叠。剔除 NT-3 基因可引起皮肤触 - 压觉感受器明显缺失; DBNF 缺乏的小鼠,其周围感觉神经元数量减少,前庭神经节严重变性。1
受体已发现神经末梢上有高亲和力和低亲和力两类 NT 受体,高亲和力受体是一类为 140 kD 的结合酪氨酸激酶的受体,包括 trk A 、 trk B 和 trk C 受体三种。 Trk A 受体对 NGF 的亲和力较高; trk B 受体对 BDNF 和 NT-4/5 的亲和力较高;而 Trk C 受体则主要与 NT-3 结合。各种受体均以二聚体的形式存在,受体激动后可促发胞浆内酪氨酸蛋白激酶的磷酸化。低亲和力受体是一种 75 kD 的膜蛋白,称为 p75 NTR 。这种受体的数量远比高亲和力受体多,约为后者的 7 倍。 P75 NTR 与 Trk A 单体形成的二杂合体能增强与 NGF 特异结合的亲和力。但由两个 p75 NTR 聚合而成的同源二聚体与 NT 结合时,则可引起相反的效应,甚至导致细胞凋亡。2
影响神经元生长的其他营养因子随着无血清培养神经元等技术的应用,在许多组织液和细胞外基质中陆续发现一些新的特异蛋白质分子,也能促进神经元的增殖、分化和存活。例如,施万细胞和星形胶质细胞产生的 睫状神经营养因子 ( ciliary neurotrophic factor, CNTF )能促进受损伤的和胚胎的脊髓神经元存活,并在治疗人类运动神经元变性疾病中有重要价值。又如, 胶质细胞源神经营养因子 ( glial cell line-derived neurotrophic factor, GDNF )在离体实验中能支持中脑多巴胺能神经元的生存,在各种帕金森病动物模型上可提高多巴胺能神经元的存活率和神经末梢的密度而改善其症状。此外,促进神经元生长的还有 白血病抑制因子( leukemia inhibitory factor, LIF )、 胰岛素样生长因子 Ⅰ ( insulin like-growth factor- Ⅰ , IGF- Ⅰ )、 转化生长因子 ( transforming growth factor, TGF )、 表皮生长因子 ( epidermal growth factor, EGF )、 成纤维细胞生长因子 ( fibroblast growth factor, FGF )和 血小板源生长因子 ( platelet-derived growth factor, PDGF )等。
神经营养因子与受体的相互作用2008年7月2日,《自然》(Nature)杂志在线发表了中国科学院生物物理所江涛课题组题为“Crystal structure of the neurotrophin-3 and p75NTR symmetrical complex” 的研究论文。该论文报道了神经营养因子3与其受体p75NTR胞外区复合物的晶体结构,研究结果揭示了神经营养因子与其受体p75NTR相互作用的方式与结构基础。3
神经营养因子是一类对神经元的发育、存活和凋亡起重要作用的蛋白质,其成员包括神经生长因子(NGF),脑源性生长因子(BDNF),神经营养因子3(NT-3),神经营养因子4(NT-4)等,这些蛋白质是治疗神经损伤等疾病的潜在药物标靶。过去的研究表明,神经营养因子有两种不同的膜蛋白受体,分别为p75NTR 受体和酪氨酸激酶受体Trk。神经营养因子通过与这两种受体的胞外区相互作用,将有关神经细胞存活和凋亡的信号传递到细胞内部,从而调控细胞的发育与凋亡。p75NTR可以与所有的神经营养因子相结合,但是p75NTR与神经营养因子的结合方式一直存在争议。此外,p75NTR与Trk之间既存在协同作用,又存在拮抗作用, 但是详细的协同调控机制仍有待进一步研究来阐明。3
神经营养因子在学术文献中的解释1、神经营养因子是指机体产生的能够促进神经细胞存活、生长、分化的一类蛋白质因子.过去一直认为神经生长因子主要在发育过程中调节神经元存活,而对成年神经元不产生作用。
2、一般将神经营养物质和上述对神经细胞存活具有调节作用的生长因子统称为神经营养因子.2 神经营养因子概述21 神经营养物质的结构及其受体神经营养物质如NGF、BDNF、NT3、NT45及NT6等。
3、营养因子这些因子又称为神经营养因子.它们是内生性肽类物质,可以通过阻止apoptosis(细胞死亡)受体P75与TGF-RI的影响而促进神经元的生长与存活。
4、在某种意义上S100蛋白可以称为一种神经营养因子,其在损伤后的神经胶质细胞中的增多,是神经组织修复在分子水平的表现兔脑出血灶周围脑组织GFAP及S-100的表达。
5、这类物质被称为神经营养因子。包括神经生长因子(nevergrowthfactorNGF)脑源性神经营养因子(brainderivedneurotrophicfactorBDNF)神经营养因子-3(neurotrophin-3NT-3)神经营养因子-45(NT-45)等。
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国