

神经传导,即神经冲动的传导过程,是电化学的过程,是在神经纤维上顺序发生的电化学变化。神经受到刺激时,细胞膜的透性发生急剧变化。用同位素标记的离子做试验证明,神经纤维在受到刺激(如电刺激)时,Na+的流入量比未受刺激时增加20倍,同时K+的流出量也增加9倍,所以神经冲动是伴随着Na+大量流入和K+的大量流出而发生的。
传导过程神经冲动的传导过程可概括为:①刺激引起神经纤维膜透性发生变化,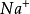 大量从膜外流入,从而引起膜电位的逆转,从原来的外正内负变为外负内正,这就是动作电位,动作电位的顺序传播即是神经冲动的传导;②纤维内的K离子向外渗出,从而使膜恢复了极化状态;③
大量从膜外流入,从而引起膜电位的逆转,从原来的外正内负变为外负内正,这就是动作电位,动作电位的顺序传播即是神经冲动的传导;②纤维内的K离子向外渗出,从而使膜恢复了极化状态;③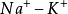 泵的主动运输使膜内的Na+流出,使膜外的K+流入,由于Na离子:K离子的主动运输量是3:2,即流出的Na离子多,流入的K离子少,也由于膜内存在着不能渗出的有机物负离子,使膜的外正内负的静息电位和Na离子、K离子的正常分布得到恢复。
泵的主动运输使膜内的Na+流出,使膜外的K+流入,由于Na离子:K离子的主动运输量是3:2,即流出的Na离子多,流入的K离子少,也由于膜内存在着不能渗出的有机物负离子,使膜的外正内负的静息电位和Na离子、K离子的正常分布得到恢复。
静息电位静息电位(Resting Potential,RP)是指细胞未受刺激时,存在于细胞膜内外两侧的外正内负的电位差。它是一切生物电产生和变化的基础。当一对测量微电极都处于膜外时,电极间没有电位差。在一个微电极尖端刺入膜内的一瞬间,示波器上会显示出突然的电位改变,这表明两个电极间存在电位差,即细胞膜两侧存在电位差,膜内的电位较膜外低。该电位在安静状态始终保持不变,因此称为静息电位。几乎所有的动植物细胞的静息电位膜内均较膜外低,若规定膜外电位为零,则膜内电位即为负值。大多数细胞的静息电位在-10~100mV之间。
细胞膜两侧的电位差在某些情况下会发生变动,使细胞膜处于不同的电位状态。细胞安静时膜两侧内负外正的状态称为膜的极化状态。当膜电位向膜内负值增大方向变化时,称为超极化;相反,膜电位向膜内负值减小方向变化,称为去极化;去极化近一步加剧,膜内电位变为正值,而膜外电位变为负值,则称为反极化;细胞受到刺激后先发生去极化,再向膜内为负的静息电位水平恢复,称为膜的复极化。
静息电位是一种稳定的直流电位,但各种细胞的数值不同。哺乳动物的神经细胞的静息电位为-70mV(即膜内比膜外电位低70mV),骨骼肌细胞为-90mV,人的红细胞为-10mV。
细胞静息时在膜两侧存在电位差的原因:①细胞膜两侧各种离子浓度分布不均;②在不同状态下,细胞膜对各种离子的通透性不同。
枪乌贼轴突膜内外主要离子分布:
|| ||
细胞膜两侧的离子呈不均衡分布,膜内的钾离子高于膜外,膜内的钠离子和氯离子低于膜外,即胞内为高钾、低钠、低氯的环境。此外,有机阴离子仅存在于细胞内。在安静状态下,细胞膜对钾离子通透性大,对钠离子通透性很小,仅为钾离子通透性的1/100~1/50,而对氯离子则几乎没有通透性。因此,静息期主要的离子流为钾离子外流。钾离子外流导致正电荷向外转移,其结果导致细胞内的正电荷减少而细胞外正电荷增多,从而形成细胞膜外侧电位高而细胞膜内侧电位低的电位差。可见,钾离子外流是静息电位形成的基础,推动钾离子外流的动力是膜内外钾离子浓度差。
钾离子外流并不能无限制地进行下去,因为随着钾离子顺浓度差外流,它所形成的内负外正的电场力会阻止带正电荷的钾离子继续外流。当浓度差形成的促使钾离子外流的力与阻止钾离子外流的电场力达到平衡时,钾离子的净移动就会等于零。此时,细胞膜两侧稳定的电位差称为钾离子的平衡电位。
根据物理化学能斯特公式,只要知道细胞膜两侧钾离子的浓度差,就可计算出钾离子的平衡电位。如果人工改变细胞膜外钾离子的浓度,当浓度增高时测得的静息电位值减小,当浓度降低时测得的静息电位值增大,其变化与根据能特斯公式计算所得的预期值基本一致。科学家注意到根据公式计算出钾离子平衡电位还是与实际测量出的静息电位有很小的一些差别的,测定值总是比计算值负得少。这是由于膜对钠离子和氯离子也有很小的通透性,它们的经膜扩散(主要指钠离子的内移),可以抵销一部分由钾离子外移造成的电位差数值。
静息状态下钾离子的外流是构成静息电位的主要因素。一般细胞内钾离子的浓度变化非常小,因此造成细胞内外钾离子浓度差变动的主要因素是细胞外的钾离子浓度。如果细胞外钾离子浓度增高,可使细胞内外的钾离子浓度差减小,从而使钾离子向外扩散的动力减弱,钾离子外流减少,结果是静息电位减小。反之,则使静息电位增高。这个实验也进一步说明,形成静息电位的主要离子就是钾离子。
插入膜内的是尖端直径
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国