

生化意义
乙酰辅酶A是人体内重要的化学物质。首先,丙酮酸氧化脱羧,脂酸的β-氧化的产物。同时,它是脂酸合成,胆固醇合成和酮体生成的碳来源。三大营养物质的彻底氧化殊途同归,都会生成乙酰辅酶A以进入三羧酸循环。
乙酰辅酶A乙酰辅酶A是能源物质代谢的重要中间代谢产物,在体内能源物质代谢中是一个枢纽性的物质。糖、脂肪、蛋白质三大营养物质通过乙酰辅酶A汇聚成一条共同的代谢通路——三羧酸循环和氧化磷酸化,经过这条通路彻底氧化生成二氧化碳和水,释放能量用以ATP的合成。乙酰辅酶A是合成脂肪酸、酮体等能源物质的前体物质,也是合成胆固醇及其衍生物等生理活性物质的前体物质。
分解代谢糖是多羟基醛和多羟基酮及其衍生物的总称。人体最重要的单糖是葡萄糖(glucose),葡萄糖是糖在体内的运输形式;人体最重要的多糖是糖原,糖原是葡萄糖在体内的储存形式;食物中的多糖主要是淀粉,淀粉由淀粉酶水解为葡萄糖后才能吸收,经血液运往全身各组织被利用或储存。糖的主要生理功能是氧化供能,每克糖彻底氧化可释能16.7 kJ(4kcal),一般由糖氧化供给的能量约占人体所需总能量的50%~70%。
【糖的有氧氧化】葡萄糖→丙酮酸→乙酰辅酶A→CO2+H2O。此过程在只能有线粒体的细胞中进行,并且必须要有氧气供应。糖的有氧氧化是机体获得ATP的主要途径,1分子葡萄糖彻底氧化为二氧化碳和水可合成30或32分子ATP(过去的理论值为36或38分子ATP)。
【糖的无氧氧化】葡萄糖→丙酮酸→乳酸。在细胞无线粒体或缺乏氧气时进行,1分子葡萄糖氧化产生2分子乳酸,净合成2分子ATP。此过程产生的乳酸如果积累过多会导致乳酸酸中毒。
【糖的磷酸戊糖途径】葡萄糖→5-磷酸核糖、NADPH。此过程的产物5-磷酸核糖是合成核苷的原料之一,NADPH是细胞内良好的还原剂,为加氢反应提供氢。
【糖原合成】葡萄糖→肝糖原、肌糖原。糖原是机体糖的贮存形式,但由于糖原的贮存需要水的存在,因此贮存量较小,也正因为糖原亲水,所以糖原的利用速度比脂肪快。
【糖转化为脂肪】葡萄糖→乙酰辅酶A→脂肪酸→脂肪。这是糖转化为脂肪的途径,脂肪是机体高度还原的能源贮存形式,疏水,可以大量贮存,但利用速度较慢。1
糖酵解葡萄糖或糖原的葡萄糖单位通过糖酵解途径分解为丙酮酸,这个过程称为糖的无氧分解。由于此过程与酵母菌使糖生醇发酵的过程基本相似,故又称糖酵解(图5-1-3)。反应在胞液中进行,不需要氧气。
糖酵解的反应过程可分两个阶段:①活化吸能阶段,通过消耗2分子ATP使1分子葡萄糖裂解为2分子3碳糖。②3碳糖氧化释放能量阶段,产生2分子丙酮酸、2分子NADH和4分子ATP。糖酵解过程净产生ATP 2分子(图5-1-4)。
在糖酵解进行过程中,有三种酶催化的反应不可逆,这三个酶称为关键酶,它们使糖酵解由葡萄糖向丙酮酸方向进行。
【己糖激酶】或肝中【葡萄糖激酶】催化葡萄糖磷酸化生成6-磷酸葡萄糖,由ATP提供能量和磷酸基团。这一步反应不仅活化了葡萄糖,使其能进入各种代谢途径,还能捕获进入细胞内的葡萄糖,使之不再透出细胞膜。反应不可逆,反应过程中消耗1分子ATP。己糖激酶或葡萄糖激酶是糖酵解途径的第一个限速酶。
【磷酸果糖激酶-1】催化6-磷酸果糖转变为1,6-二磷酸果糖,这是酵解途径中的第二个磷酸化反应,需要ATP和Mg,反应不可逆。磷酸果糖激酶-1是糖酵解过程中最重要的限速酶。此酶为变构酶。柠檬酸、ATP为变构抑制剂,ADP、AMP和 F-1,6-BP等为变构激活剂。胰岛素诱导其生成。
【丙酮酸激酶】催化磷酸烯醇式丙酮酸转变为丙酮酸,磷酸烯醇式丙酮酸的高能磷酸键在催化下转移给ADP生成ATP,自身生成烯醇式丙酮酸后自发转变为丙酮酸。反应不可逆。是糖酵解途径中第二个以底物水平磷酸化方式生成ATP的反应。丙酮酸激酶是糖酵解途径中的又一个限速酶,具有别构酶特性,ATP是其别构抑制剂,ADP是别构激活剂。
在糖酵解过程中有2步反应生成ATP,其一是在磷酸甘油酸激酶催化下将1,3-二磷酸甘油酸分子上的1个高能磷酸键转移给ADP生成ATP;另1个是丙酮酸激酶催化使磷酸烯醇式丙酮酸的高能磷酸键转移给ADP生成ATP。这两步反应的共同点是底物分子都具有高能键,底物分子的高能键转移给ADP生成ATP的方式称为【底物水平磷酸化】。底物水平磷酸化是ATP的生成方式之一,另一种ATP的生成方式是氧化过程中脱下的氢(以NADH和FADH2形式存在)在线粒体中氧化成水的过程中,释放的能量推动ADP与磷酸合成为ATP,这种方式称为【氧化磷酸化】(见本章第二节)。
丙酮酸的去路糖酵解过程的产物丙酮酸有多种分支去路
1.生成乙酰辅酶A:丙酮酸在有氧气和线粒体存在时进入线粒体,经丙酮酸脱氢酶复合体(表5-1-2)催化氧化脱羧产生NADH、CO2和乙酰辅酶A,乙酰辅酶A进入三羧酸循环和氧化磷酸化彻底氧化为CO2和H2O,释放的能量在此过程中可产生大量ATP。这是糖的有氧氧化过程。糖的有氧氧化是机体获得ATP的主要途径。
丙酮酸生成乙酰辅酶A的反应是糖有氧氧化过程中重要的不可逆反应。丙酮酸脱氢产生NADH+H,释放的自由能则贮于乙酰辅酶A中。乙酰辅酶A可参与多种代谢途径。
丙酮酸脱氢酶系的多种辅酶中均含有维生素,TPP中含有维生素B1,辅酶A(HSCoA)中含有泛酸,FAD含有维生素B2,NAD含尼克酰胺(维生素PP)。所以,当这些维生素缺乏,特别是维生素B1缺乏时,丙酮酸及乳酸堆积,能量生成减少,可发生多发性末梢神经炎,严重时可引起典型脚气病。
2.丙酮酸在无氧或无线粒体条件下加氢还原为乳酸。糖酵解过程生成的产物有3个:NADH、ATP和丙酮酸。NADH、ATP的生成必将导致底物NAD和ADP的显著减少,而这两种底物的减少将严重抑制糖酵解的继续进行。ATP在体内会很快被消耗而生成ADP和磷酸,因此ATP的抑制作用几乎可以忽略不计。NADH在有氧气存在的条件下在线粒体中被氧化为水而重新生成NAD,但在无氧或无线粒体的细胞中是无法进行这个过程的,因此NAD的减少和NADH的增多在无氧或无线粒体的细胞中对糖酵解的抑制非常显著。在这些细胞中解决的办法是,产物丙酮酸作为受氢体将NADH的氢接受重新生成NAD, 丙酮酸加氢还原为乳酸。
乳酸的生成使NAD再生,能在一定时间内暂时解除糖酵解的抑制,但是如果乳酸进一步增多,乳酸的抑制作用将增强,最后糖酵解被完全抑制。同时乳酸解离产生的H也增多,体液pH下降。这些综合结果被称为【乳酸酸中毒】。在缺氧和剧烈运动时最容易产生乳酸中毒现象。乳酸中毒的解除需依赖氧气的充分供应,此时,乳酸可脱氢生成丙酮酸通过有氧氧化代谢或进入肝脏进行糖异生。
红细胞缺乏线粒体,因此,红细胞只能依赖糖的无氧氧化(酵解)获得能量,所释放的乳酸经血液循环至肝脏代谢(糖异生)。某些组织细胞如视网膜、睾丸、白细胞、肿瘤细胞等,即使在有氧条件下仍以糖酵解为其主要供能方式。
机体在缺氧情况下,尤其在剧烈运动时肌肉的氧分得不到足够供应(尽管此时气喘吁吁),糖的无氧氧化(葡萄糖→乳酸)是机体获得能量的一种有效方式,但无法维持很长时间,如果导致严重的乳酸中毒,又不能恢复氧气供应,糖酵解被完全抑制,ATP消耗不能再生,生命过程将终止。
3.丙酮酸经转氨基作用生成丙氨酸,作为蛋白质合成的原料。
4.在植物和酵母菌细胞内,无氧情况下丙酮酸脱羧产生乙醛,乙醛由NADH还原为乙醇(乙醇发酵)。乙醇发酵有很大的经济意义,在发面、制作面包和馒头,以及酿酒工业中起着关键性的作用。在酿醋工业上,微生物也是先在不需氧条件下形成乙醛而后在有氧条件下氧化为乙酸(醋酸)。
糖酵解的调节正常生理条件下,人体内的各种代谢过程受到严格而精细的调节,以保持内环境稳定,适应机体生理活动的需要。这种调节控制主要是通过改变酶的活性来实现的。己糖激酶(葡萄糖激酶)、磷酸果糖激酶-1、丙酮酸激酶是糖酵解的关键酶,它们的活性大小,直接影响着整个代谢途径的速度和方向,其中以磷酸果糖激酶-1最为重要。
1.激素的调节 胰岛素可诱导GK、PFK-1、PK的合成,因而使糖酵解过程增强。
2.代谢物对限速酶的变构调节磷酸果糖激酶-1(PFK-1)是三个限速酶中催化效率最低的,故而是糖酵解途径中最重要的调节点。该酶分子为四聚体。分子中不仅具有与底物结合的部位,还具有与变构激活剂和变构抑制剂结合的部位。F-1,6-BP、ADP、AMP等是其变构激活剂,而ATP、柠檬酸等为其变构抑制剂。在这些代谢物的共同调节下,机体可根据能量需求调整糖分解速度。当细胞内能量消耗增多,ATP浓度降低,AMP、ADP浓度增加,则磷酸果糖激酶-1被激活,糖分解速度加快,使ATP生成量增加;当细胞内有足够的ATP储备时,ATP浓度增加,AMP、ADP浓度下降,磷酸果糖激酶-1被抑制,糖分解速度减慢,减少ATP生成量,避免能量的浪费;当饥饿时,机体动员储存脂肪分解氧化,生成大量乙酰CoA,乙酰CoA可与草酰乙酸缩合成柠檬酸,抑制磷酸果糖激酶-1的活性,从而减少糖的分解,以维持饥饿状态下血糖浓度。
糖酵解的分支磷酸戊糖途径(pentose phosphate pathway)由6-磷酸葡萄糖开始,全过程可分为二个阶段:第一阶段是6-磷酸葡萄糖脱氢氧化生成NADPH+H 、CO2和5-磷酸核糖。第二阶段为一系列基团转移反应。
1.反应过程(图5-1-6)
(1)5-磷酸核糖生成 6-磷酸葡萄糖在6-磷酸葡萄糖脱氢酶和6-磷酸葡萄糖酸脱氢酶相继催化下,经2次脱氢和1次脱羧,生成2分子NADPH+H 和1分子CO2后生成5-磷酸核酮糖,5-磷酸核酮糖经异构酶催化转变为5-磷酸核糖。
(2)基团移换反应 此阶段由4分子5-磷酸木酮糖和2分子5-磷酸核糖在转酮基酶、转醛基酶催化下,通过一系列反应,最后生成4分子6-磷酸果糖和2 分子3-磷酸甘油醛。2分子3-磷酸甘油醛可缩合成1分子6-磷酸果糖。综上所述,1分子6-磷酸葡萄糖经磷酸戊糖途径氧化,需5 分子6-磷酸葡萄糖伴行,最后又生成5分子6-磷酸葡萄糖,实际消耗1分子6-磷酸葡萄糖。
磷酸戊糖途径中的限速酶是6-磷酸葡萄糖脱氢酶,此酶活性受NADPH+H浓度影响,NADPH+H 浓度增高时抑制该酶活性,因此磷酸戊糖途径的代谢速度主要受细胞内NADPH+H 需求量的调节。
2.磷酸戊糖途径的生理意义
磷酸戊糖途径的主要生理意义是产生5-磷酸核糖和NADPH+H。
(1)生成5-磷酸核糖(R-5-P):磷酸戊糖途径是体内利用葡萄糖生成5-磷酸核糖的唯一途径。5-磷酸核糖是合成核酸和核苷酸辅酶的重要原料。对于缺乏6-磷酸葡萄糖脱氢酶的组织如肌肉,也可利用糖酵解中间产物3-磷酸甘油醛和6-磷酸果糖经转酮基酶和转醛基酶催化的逆反应生成。故损伤后修复的再生组织、更新旺盛的组织,此途径都比较活跃。
(2)生成NADPH+H :①NADPH+H是体内许多合成代谢中氢原子的供体,如脂肪酸、胆固醇和类固醇激素等化合物的合成,都需要大量NADPH,因此在脂肪、固醇类化合物合成旺盛的组织,如肝脏、哺乳期乳腺、脂肪组织、肾上腺皮质及睾丸等组织中,磷酸戊糖途径特别活跃。②NADPH+H是谷胱甘肽(GSH)还原酶的辅酶,对于维持细胞中谷胱甘肽于还原状态起重要作用。GSH是细胞中重要的抗氧化物质,有清除H2O2和过氧化物,保护细胞中含巯基的酶和蛋白质免遭氧化破坏的作用,以维持细胞结构和功能的完整。红细胞中如发生H2O2和过氧化物的积累,将使红细胞的寿命缩短并增加血红蛋白氧化为高铁血红蛋白的速率,后者没有运氧功能。遗传性6-磷酸葡萄糖脱氢酶缺陷的患者,磷酸戊糖途径不能正常进行,NADPH+H缺乏,GSH含量减少,常在进食蚕豆或使用某些药物后诱发急性溶血性黄疸。③NADPH+H是加单氧酶系的组成成分,参与激素、药物、毒物的生物转化过程。
(3)中间产物3-磷酸甘油醛:3-磷酸甘油醛是三种代谢途径的枢纽。如果磷酸戊糖途径受阻,3-磷酸甘油醛则可进入糖的无氧分解或糖的有氧分解途径;反之,若用碘乙酸抑制3-磷酸甘油醛脱氢酶,使糖的无氧分解和有氧分解不能进行,则3-磷酸甘油醛可进入磷酸戊糖途径。磷酸戊糖途径在整个代谢过程中没有氧的掺入,但可使葡萄糖降解,这在种子萌发的初期作用很大。植物染病或受伤时,磷酸戊糖途径增强,所以它与植物的抗病能力有一定关系。
(4)与植物光合作用有关:磷酸戊糖途径产生的三碳糖、五碳糖、七碳糖都是植物光合作用的中间产物,有的反应是光合作用与卡尔文循环中某些反应的相应逆反应,而且它与卡尔文循环还有一些相同的酶。
氧化分解脂肪(三脂酰甘油或甘油三酯)在体内主要功能是氧化分解,为机体提供生命活动所需要的能量。储存于脂肪组织中的三脂酰甘油 (triglyceride),被脂肪酶逐步水解为游离脂肪酸(free fatty acid,FFA)及甘油释放入血,供给全身各组织氧化利用的过程,称为三脂酰甘油动员。脂肪组织中含有的脂肪酶,一种是对激素敏感的三脂酰甘油脂肪酶,称激素敏感脂肪酶(hormone-sensitive triglyceride lipase,HSL) ,也是三脂酰甘油动员的限速酶。另外还有二脂酰甘油脂肪酶和一脂酰甘油脂肪酶,这两种酶活性较高,对激素不敏感。胰高血糖素、肾上腺素、去甲肾上腺素等能激活三脂酰甘油脂肪酶,促进脂肪分解,故称为脂解激素;胰岛素的作用则与之相反,使三脂酰甘油脂肪酶活性降低,拮抗脂解激素的脂解作用,故称抗脂解激素。一分子脂肪被脂肪酶水解为1分子甘油和3分子脂肪酸后,分别进入各自的代谢途径。
甘油的氧化分解甘油主要由心、肝、骨骼肌等组织摄取利用,在细胞内经甘油激酶(glycerokinase)的作用,生成α-磷酸甘油(3-磷酸甘油),后者在α-磷酸甘油脱氢酶的催化下生成磷酸二羟丙酮,磷酸二羟丙酮可循糖代谢途径氧化分解释放能量,1分子甘油彻底氧化可净生成17.5~19.5分子ATP。也可以在肝脏循糖异生途径转变为糖原和葡萄糖。
脂肪酸的氧化在氧供给充足的条件下,脂肪酸可在体内分解成二氧化碳和水,释出大量能量。除脑组织和成熟红细胞外,大多数组织均能氧化脂肪酸,但以肝及肌肉组织最活跃。
1.脂肪酸的活化——脂酰CoA的生成
脂肪酸的活化反应在胞液中进行,脂肪酸在脂酰CoA合成酶(acyl-CoA synthetase)催化下,在ATP、CoA-SH、Mg存在的条件下,活化为脂酰CoA。脂酰CoA含有高能硫酯键,而且水溶性增大,使脂酰基的代谢活性明显增加。分子中的CoA是脂酰基的载体。由于反应过程中生成的焦磷酸(PPi),迅速被细胞内的焦磷酸酶水解,阻止了逆向反应的发生,因此1分子脂肪酸活化成脂酰CoA,实际上消耗了2个高能磷酸键。
2.脂酰CoA进入线粒体
脂肪酸的活化在胞液中进行,而催化脂肪酸氧化分解的酶系存在于线粒体基质,因此活化的脂酰CoA必须进入线粒体才能分解。脂酰CoA不能直接透过线粒体内膜,其脂酰基需经肉毒碱(carnitine即3-羟-4-三甲氨基丁酸)转运才能进入基质。线粒体内膜的两侧存在着肉毒碱脂酰转移酶Ⅰ及Ⅱ,在位于线粒体内膜外侧面的酶Ⅰ的催化下,脂酰CoA转化为脂肪酰肉毒碱,而移到膜内侧,进入膜内侧的脂肪酰肉毒碱又经酶Ⅱ的催化而重新转变成脂酰CoA,并释放出肉毒碱。肉毒碱脂酰转移酶Ⅰ是限速酶,脂酰CoA进入线粒体是脂肪酸氧化的限速步骤,当饥饿、高脂低糖膳食或糖尿病时,体内糖利用发生障碍,需要脂肪酸供能,这时肉毒碱脂酰转移酶Ⅰ活性增加,脂肪酸氧化增强。
3.脂酰基的β-氧化
脂酰基进入线粒体基质后,从脂酰基的β-碳原子开始,经过脱氢、加水、再脱氢及硫解四步连续的反应,脂酰基断裂产生1分子乙酰CoA和1分子比原来少两个碳原子的脂酰CoA(图5-1-9)。催化这些反应的酶彼此结合形成多酶复合体,称脂肪酸氧化酶系。
(1)脱氢:脂酰CoA在脂酰CoA脱氢酶的催化下,α、β碳原子各脱去一个氢原子,生成α,β-烯脂酰CoA,脱下的2H由FAD接受生成FADH2。一分子FADH2进入呼吸链通过氧化磷酸化产生1.5分子ATP(过去的理论值为2分子ATP,详见氧化磷酸化一节)。
(2)加水:在水化酶催化下,烯脂酰CoA加水生成β-羟脂酰CoA。
(3)再脱氢:在β-羟脂酰CoA脱氢酶催化下,β-羟脂酰CoA脱下2H,生成β-酮脂酰CoA,脱下的2H由NAD接受,生成NADH+H。一分子NADH+H进入呼吸链通过氧化磷酸化产生2.5分子ATP(过去的理论值为3分子ATP)。
(4)硫解(加CoASH分解):β-酮脂酰CoA在硫解酶的催化下,加入CoASH使α、β碳原子之间化学键断裂,生成1分子乙酰CoA和1分子比原来少两个碳原子的脂酰CoA。
以上生成的比原来少2个碳原子的脂酰CoA可再进行脱氢、加水、再脱氢及硫解反应,如此反复进行,直至最后生成丁酰CoA,后者再进行一次β-氧化,即完成脂肪酸的β-氧化。
4.乙酰CoA的去路:脂肪酸β-氧化的终产物是乙酰CoA,其进一步的代谢变化可进入三羧酸循环彻底氧化为二氧化碳和水,也可转变为其它物质。
5.脂肪酸彻底氧化时能量的释放和利用
脂肪酸氧化是体内能量的重要来源。以16个碳原子的软脂酸为例:一分子软脂酰CoA需经7次β-氧化,生成8分子乙酰CoA。因此1分子软脂酸彻底氧化共生成(7×1.5)+(7×2.5)+(8×10)=108分子ATP。减去软脂酸活化时消耗的两个高能磷酸键,净生成106分子ATP。
生成利用脂肪酸在肝外组织(如心肌、骨骼肌等)经β-氧化生成的乙酰CoA,能彻底氧化生成二氧化碳和水,而在肝细胞中因为具有活性较强的合成酮体的酶系,β-氧化反应生成的乙酰CoA,大多转变为乙酰乙酸(acetoacetate),β-羟丁酸(β-hydroxybutyrate)和丙酮(acetone),这三种中间产物统称酮体(ketone bodies)。由于肝内缺乏氧化利用酮体的酶系,所以酮体不能在肝内氧化,必须透过细胞膜进入血液循环运输到肝外组织才能进一步氧化分解供能。
酮体的生成酮体生成的部位是在肝细胞线粒体内。脂肪酸β-氧化生成的乙酰CoA是合成酮体的原料。其合成过程分三步进行(图5-1-10)。
1.两分子乙酰CoA在硫解酶(thiolase)催化下缩合成1分子乙酰乙酰CoA。
2.乙酰乙酰CoA再与1分子乙酰CoA缩合成β-羟-β-甲基戊二酸单酰CoA(HMG-CoA),催化这一反应的酶为HMG-CoA合成酶,是酮体合成的限速酶。
3.HMG-CoA经裂解酶催化分解成乙酰乙酸和乙酰CoA。乙酰乙酸加氢还原成β-羟丁酸,少量乙酰乙酸自发脱羧生成丙酮。酮体总量中约70%为β-羟丁酸,30%为乙酰乙酸,丙酮只占少量,丙酮可通过肾和肺排出。
酮体的利用肝外组织(心肌、骨骼肌、大脑)中有活性很强的利用酮体的酶。乙酰乙酸在乙酰乙酸硫激酶或琥珀酰CoA转硫酶催化下,转变为乙酰乙酰CoA,然后再被硫解酶分解为两分子乙酰CoA,乙酰CoA进入三羧酸循环彻底氧化。可见肝内生酮肝外用是脂肪酸在肝中氧化的一个代谢特点。
生理意义酮体是脂肪酸在肝脏氧化的正常中间产物,是肝脏为肝外组织提供能源物质的一种形式,酮体分子小、溶于水,便于通过血液运输,也易于通过血脑屏障及肌肉等组织的毛细血管壁,是肌肉,尤其是脑组织的重要能源。脑组织不能氧化脂肪酸,却能利用酮体。长期饥饿、糖供应不足时,酮体可以代替葡萄糖,成为脑组织的主要能源物质。
正常情况下血中仅含少量酮体。但在饥饿、妊娠呕吐及糖尿病时,三脂酰甘油动员加强,肝中酮体生成过多,超出肝外组织利用的能力,可引起血中酮体升高,尿中出现酮体,即酮血症和酮尿症,可导致酮症酸中毒,严重者危及生命。
分解代谢氨基酸的主要功用是作为蛋白质合成的原料;其次可合成其它含氮物质(如嘌呤、嘧啶等);过多的氨基酸在体内不能贮存,这部分氨基酸可通过各种代谢方式先转变为三羧酸循环的中间产物,然后经三羧酸循环彻底氧化为CO2和H2O,也可通过糖异生作用转变为葡萄糖,还可转变为脂肪贮存。各种氨基酸具有共同的结构特点,故有共同的代谢途径,但不同的氨基酸由于结构的差异也有不同的代谢方式。
三羧酸循环右图显示氨基酸进入三羧酸循环的方式。从图中可以看出,10种氨基酸最后分解产生乙酰CoA ;5种氨基酸转变成三羧酸循环的中间产物α-酮戊二酸;4种氨基酸转变成琥珀酰CoA;2种氨基酸转变为延胡索酸;2种氨基酸转变为草酰乙酸。然后可经三羧酸循环进一步彻底氧化为CO2和H2O。2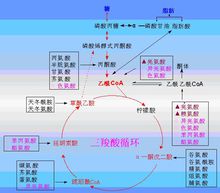
氨基酸也可异生为糖或生成酮体。凡是能转变为丙酮酸、草酰乙酸、琥珀酰CoA、α-酮戊二酸和延胡索酸的氨基酸称为生糖氨基酸,这是因为三羧酸循环的中间产物和丙酮酸能转变为磷酸烯醇式丙酮酸,然后很容易循糖异生途径异生为糖。凡是能分解为乙酰CoA和乙酰乙酰CoA的氨基酸称为生酮氨基酸,因为乙酰CoA或乙酰乙酰CoA亦可用于合成脂肪。20种氨基酸中,只有亮氨酸和赖氨酸是唯一生酮的氨基酸;异亮氨酸、苯丙氨酸、色氨酸和酪氨酸是生酮兼生糖氨基酸;剩余14种是生糖氨基酸。
脱氨基作用氨基酸脱氨基作用是氨基酸分解代谢的最主要反应。体内大多数组织细胞均可进行。氨基酸可通过多种方式脱去氨基,如转氨基、氧化脱氨基、联合脱氨基等,其中以联合脱氨基最为重要。
氨基酸脱氨基的产物为α-酮酸和氨.
1.转氨基作用
大多数氨基酸在进行分解代谢之初,首先通过转氨基作用将α-氨基转移给α-酮戊二酸,使其形成谷氨酸和相应的α-酮酸(α-ketoacid)。
转氨基作用是氨基酸在氨基转移酶(aminotransferase)或称转氨酶(transaminase)催化下,可逆地把α-氨基酸的氨基转移给α-酮戊二酸,使α-氨基酸转变为相应的α-酮酸,而原来的α-酮戊二酸接受氨基转变成相应的谷氨酸。可见,转氨基作用既是氨基酸的分解代谢过程,又是某些非必需氨基酸合成的重要途径。转氨酶的辅酶是维生素B6的磷酸酯磷酸吡哆醛。体内大多数氨基酸均能进行转氨基反应,转氨酶的种类很多,专一性强,分布也最广。以丙氨酸氨基转移酶(alanine transaminase,ALT;又称谷丙转氨酶,GPT)以及天冬氨酸氨基转移酶(aspartate transaminase,AST;又称谷草转氨酶,GOT)最重要,前者在肝细胞含量最高,后者在心肌细胞含量最高。正常情况下它们在血清中含量都很低,当肝细胞或心肌受损时血清中含量增高,故可用于临床上肝脏或心肌疾病的辅助诊断。
2.谷氨酸的氧化脱氨基作用
通过以上转氨基作用生成的谷氨酸由谷氨酸脱氢酶(glutamate dehydrogenase)催化,脱氢的同时又脱去氨基的反应,称为氧化脱氨基作用。在体内氨基酸氧化酶种类很多,其中以谷氨酸脱氢酶的作用最重要。谷氨酸脱氢酶是以NAD或NADP为辅酶的不需氧脱氢酶,催化谷氨酸脱氢生成亚谷氨酸,然后水解生成α-酮戊二酸和NH3(图5-1-14)。谷氨酸脱氢酶广泛存在于肝、肾及脑中,反应可逆,通过还原氨基化作用,α-酮戊二酸和氨可合成谷氨酸,因此,它不仅在氨基酸的分解中起作用,而且在非必需氨基酸合成中也起着重要作用。
3.联合脱氨基作用
联合脱氨基作用是体内脱氨基的主要方式,生物体内存在二种联合脱氨基方式。
(1)转氨酶与谷氨酸脱氢酶的联合脱氨基作用:①氨基酸首先与α-酮戊二酸进行转氨基反应,生成相应的α酮酸和谷氨酸,②谷氨酸在谷氨酸脱氢酶作用下脱去氨基生成α-酮戊二酸。全过程可逆,通过其逆过程可以合成新的非必需氨基酸。此过程主要存在于肝、肾和脑组织中,心肌和骨骼肌中不能进行,因为心肌和骨骼肌中谷氨酸脱氢酶活性低。
(2)嘌呤核苷酸循环形式的联合脱氨基作用
肌肉组织中-谷氨酸脱氢酶活性不高,难以进行上述联合脱氨基作用,在肌肉中氨基酸是通过嘌呤核苷酸循环脱去氨基的。过程为:①α-氨基酸首先与α-酮戊二酸转氨基作用生成谷氨酸,后者再与草酰乙酸转氨基反应,生成天冬氨酸;②天冬氨酸与次黄嘌呤核苷酸(IMP)由腺苷酸代琥珀酸合成酶催化生成腺苷酸代琥珀酸;③腺苷酸代琥珀酸裂解生成腺苷酸(AMP)和延胡索酸;④AMP在腺苷酸脱氨酶(此酶在肌肉组织中活性较强)催化下脱去氨基生成IMP,完成氨基酸的脱氨基作用。IMP可以再参加循环,延胡索酸经三羧酸循环转变为草酰乙酸后再次参加转氨反应。
氨基酸脱氨基作用的终产物是α-酮酸和氨。它们将分别进入各自的代谢途径。
α-酮酸代谢氨基酸脱氨后生成的 α-酮酸可进一步代谢。主要有以下三方面:
1.经氨基化生成非必需氨基酸
实验证明人体不能合成赖、异亮、苯丙、亮、色、缬、苏、蛋等8种氨基酸相对应的α-酮酸,因而这些氨基酸不能在体内合成,必须从食物摄取,称为营养必需氨基酸。其它十二种氨基酸则称为营养非必需氨基酸,所谓非必需氨基酸并不是它们在代谢中的作用不重要,而是可以在人体合成,主要通过联合脱氨基作用的逆反应生成,故食物不给与一般不会引起缺乏。
2.转变成糖或脂肪
如前述,在体内可以转变成糖的氨基酸称为生糖氨基酸(glucogenic amino acid),能转变为酮体者称为生酮氨基酸(ketogenic amino acid),二者兼有的则称为生糖兼生酮氨基酸(glucogenic and ketogenic amino acid)。
3.氧化供能 不同的α-酮酸在体内可以通过三羧酸循环与氧化磷酸化彻底氧化,产生CO2和水,并释放出能量供生命活动的需要。
氨的代谢氨是一种剧毒物质,脑组织对氨的作用尤为敏感,需要及时处理以免在组织中堆积。正常人除门静脉血液外,血液中氨的浓度极低,一般不超过60μmol/L(0.1mg/dl)。
1.体内氨的来源
(1)氨基酸分解产生氨:氨基酸脱氨基作用是氨的主要来源;胺类物质的氧化分解也可产生氨。
(2)肠道吸收:肠道氨主要来自①肠道细菌对未被消化的蛋白质和未被吸收的氨基酸作用(称腐败作用)产生的氨;②血中尿素扩散入肠管后在肠道细菌尿素酶作用下水解产生的氨。NH3比NH4容易穿过细胞膜而被吸收,在碱性环境中,NH4转变为NH3,所以肠管pH偏碱时,氨的吸收增加。临床上对高血氨病人采用酸性透析液做结肠透析而不用碱性肥皂水灌肠就是这个道理。肠道每日产氨约有4g,腐败作用增强时,氨的产生更多。
(3)肾脏产生:谷氨酰胺在肾远曲小管上皮细胞谷氨酰胺酶的催化下,水解生成谷氨酸和NH3,NH3分泌到肾小管腔与尿中H结合生成NH4由尿排出。碱性尿液不利NH3的分泌,NH3被吸收入血,成为血氨的另一个来源。故肝硬化腹水者不宜使用碱性利尿药以防血氨升高。
2.氨的转运
氨是有毒物质,各组织中产生的氨必须以无毒形式经血液运输至肝合成尿素或以铵盐形式随尿排出。氨在血液中有两种运输形式:
(1)丙氨酸运氨作用:主要将肌肉氨基酸脱下的氨经血液运输到肝。过程为:①肌肉中的氨基酸经转氨基作用将氨基转移给丙酮酸生成丙氨酸,经血液运输至肝;②在肝中,丙氨酸经联合脱氨基作用释放出氨,氨用于合成尿素,生成的丙酮酸则异生为葡萄糖;③葡萄糖经血液运送到肌肉,在肌肉活动供能的过程中又可分解为丙酮酸,再次接受氨基生成丙氨酸输送到肝脏。如此通过丙氨酸和葡萄糖的互变把氨从肌肉运输到肝脏的循环称丙氨酸-葡萄糖循环(alanine glucose cycle)。
(2)谷氨酰胺的运氨作用:氨与谷氨酸在ATP供能和谷氨酰胺合成酶催化下合成谷氨酰胺,经血液输送到肝或肾,经谷氨酰胺酶水解为谷氨酸及氨,在肝可合成尿素,在肾则以铵盐形式由尿排出。谷氨酰胺生成的意义:①肝外组织解除氨毒;②是从脑、肌肉等组织向肝或肾运输氨的主要形式;③氨的储存形式,为某些含氮化合物的合成提供原料,如嘌呤及嘧啶的合成。临床上对肝性脑病患者可服用或输入谷氨酸盐以降低血氨浓度。
3.氨的主要去路
氨在体内的主要去路是在肝内通过鸟氨酸循环(尿素循环)生成无毒的尿素,然后由肾排出体外)。鸟氨酸循环的过程可分为以下四步:
1)氨基甲酰磷酸的合成:氨由丙氨酸与谷氨酰胺转运入肝细胞线粒体在氨基甲酰磷酸合成酶Ⅰ(carbamoyl phosphate synthetaseⅠ,CPS-Ⅰ)催化下,与CO2和H2O分子结合,消耗2分子ATP,合成氨基甲酰磷酸。反应不可逆。
(2)瓜氨酸的合成:在鸟氨酸氨甲酰转移酶(ornithine carbamoyl transferase,OCT)催化下,将氨基甲酰磷酸的氨甲酰基转移至鸟氨酸的δ-NH2上生成瓜氨酸。反应不可逆。所需的鸟氨酸是由胞液经线粒体内膜上的载体转运进入线粒体的。合成的瓜氨酸又由线粒体内膜上的载体转运进入胞液。
(3)精氨酸的合成:在胞液内,瓜氨酸与天冬氨酸在精氨酸代琥珀酸合成酶(argninosuccinate synthetase)的催化下,由ATP供能合成精氨酸代琥珀酸并生成AMP+PPi,精氨酸代琥珀酸在精氨酸代琥珀酸裂解酶(argninosuccinate lyase)催化下,分解成为精氨酸和延胡索酸。在此过程中,天冬氨酸起着供给氨基的作用;生成的延胡索酸经三羧酸循环转变为草酰乙酸后可与α-氨基酸经转氨作用转变为天冬氨酸。由此可见,鸟氨酸循环与三羧酸循环可联系在一起。
(4)精氨酸水解生成尿素:精氨酸在胞液中精氨酸酶(arginase)的作用下,水解生成尿素和鸟氨酸。鸟氨酸再进入线粒体参与瓜氨酸的合成,反复循环,不断合成尿素。
尿素分子中的两个-NH2,一个由丙氨酸或谷氨酰胺转运入肝细胞线粒体的NH3,另一个由天冬氨酸提供,碳原子来自CO2,天冬氨酸和谷氨酸均是氨的载体。另外,尿素合成是耗能过程,每合成1分子尿素需消耗3分子ATP(消耗4个高能磷酸键)。
尿素主要通过肾脏排泄。如肾排泄功能障碍,必然导致血尿素增高。故临床常测定血尿素氮(blood urea nitrogen,BUN)来反映肾功能。
4.高血氨与氨中毒
正常情况下血氨浓度维持在较低水平。肝脏几乎是体内唯一能合成尿素的器官,当肝功能严重损伤时,尿素合成障碍,血氨浓度升高,称为高氨血症。一般认为,氨进入脑组织可与α-酮戊二酸结合生成谷氨酸,氨与谷氨酸再进一步结合生成谷氨酰胺。因此,脑中氨的增加,可消耗脑组织中α-酮戊二酸,导致三羧酸循环速度减弱, ATP生成减少,引起大脑功能障碍,严重时可产生昏迷,即氨中毒(肝性脑病)。
彻底氧化乙酰CoA是生化代谢中的一个枢纽性物质,如前所述,糖、脂肪、氨基酸分解代谢都能产生乙酰辅酶A;乙酰辅酶A有多种代谢去路,可以合成脂肪酸、胆固醇、酮体等,乙酰辅酶A彻底氧化释放能量的途径是三羧酸循环。通过三羧酸循环和氧化磷酸化,乙酰CoA氧化产生CO2、H2O,释放能量推动ATP合成。在营养物质产能代谢中,三羧酸循环和氧化磷酸化是释放能量最多的环节,是营养物质产能代谢和相互转化的枢纽。
三羧酸循环三羧酸循环(tricarboxylic acid cycle)是由Hans Adolf Krebs于1937年首先提出,故又称为Krebs循环(尿素循环也是Krebs提出的)。此循环是从活性二碳化合物—乙酰辅酶A和四碳草酰乙酸在线粒体内缩合成含三个羧基的柠檬酸开始,经过一系列脱氢脱羧反应,最后重新生成草酰乙酸而成为循环(图5-2-1)。此反应过程是由含有三个羧基的柠檬酸作为第一个中间产物的循环反应,故称为三羧酸循环,也称柠檬酸循环。三羧酸循环在线粒体内进行。
(一)三羧酸循环的反应过程
1.乙酰辅酶A与草酰乙酸缩合为柠檬酸
此反应为三羧酸循环的关键反应之一,是由柠檬酸合成酶催化的不可逆反应,所需能量来自乙酰CoA的高能硫酯键水解供应。
2. 柠檬酸转变为异柠檬酸
柠檬酸本身不易氧化,在顺乌头酸酶作用下,通过脱水与加水反应,使羟基由β碳原子转移到α碳原子上,生成易于脱氢氧化的异柠檬酸,为进一步的氧化脱羧反应作准备。
3.异柠檬酸氧化脱羧生成α-酮戊二酸
异柠檬酸在异柠檬酸脱氢酶的催化下,氧化脱羧生成α-酮戊二酸,反应脱下的氢由NAD接受生成NADH+H,脱羧使6碳化合物变为5碳化合物。这是三羧酸循环中第一次氧化脱羧生成CO2的反应。此反应不可逆,是三羧酸循环中的限速步骤。
4.α-酮戊二酸氧化脱羧生成琥珀酰辅酶A
这是三羧酸循环的第二次氧化脱羧反应,此反应类似于丙酮酸的氧化脱羧,也是不可逆反应,生成的琥珀酰CoA含有高能硫酯键。催化此反应的α-酮戊二酸脱氢酶复合体是三羧酸循环的限速酶,它由三个酶(α-酮戊二酸脱氢酶、琥珀酰基转移酶、二氢硫辛酸脱氢酶)和五个辅助因子(TPP、硫辛酸、HSCoA、NAD、FAD)组成(原理见图4-1-14)。
5.琥珀酰辅酶A生成琥珀酸
琥珀酰CoA在琥珀酸硫激酶的作用下,高能硫酯键水解,能量转移给GDP生成GTP和琥珀酸。生成的GTP可直接利用,也可将其高能磷酸基团转移给ADP生成ATP。这是三羧酸循环中唯一的一次底物磷酸化反应。
6.琥珀酸脱氢
琥珀酸在琥珀酸脱氢酶的催化下脱氢生成延胡索酸,脱下的氢由FAD接受生成FADH2。
7.苹果酸生成
延胡索酸在延胡索酸酶催化下,加水生成苹果酸。
8.草酰乙酸再生
苹果酸在苹果酸脱氢酶催化下脱氢生成草酰乙酸,脱下的氢由NAD接受生成NADH+H。再生的草酰乙酸可再次进入三羧酸循环用于柠檬酸的合成。
(二)三羧酸循环的深入讨论
1.三羧酸循环是在有氧的条件下,在线粒体内进行的循环反应过程。
三羧酸循环的产物有NADH+H、FADH2、ATP、CO2,这些产物对三羧酸循环的抑制效果不同。CO2经血循环至肺排出浓度降低,ATP快速消耗再生出ADP,因此在正常情况下这两种产物对三羧酸循环的抑制可以忽略不计。NADH、FADH2的受氢体主要是氧,因此在缺氧情况下NADH和FADH2无法将氢传递出去,致使NAD和FAD无法再生,三羧酸循环因此被抑制。这就是为什么三羧酸循环不消耗氧气但却只能在有氧条件下进行的原因。
2.三羧酸循环是机体主要的产能途径。
每循环一周,1分子的乙酰辅酶A被氧化,三羧酸循环直接消耗的底物是乙酰基。循环中有两次脱羧和四次脱氢反应,两次脱羧反应生成两分子CO2;四次脱氢反应,有三次由NAD接受共生成3分子NADH+H,有一次由FAD接受生成1分子FADH2(图5-2-2)。每个NADH+H经氧化磷酸化产生2.5个ATP共7.5个ATP,每个FADH2经氧化磷酸化产生1.5ATP共1.5个ATP,循环一周以此种方式可生成9分子ATP;加上一次底物磷酸化生成的GTP,三羧酸循环一周共可生成10分子ATP。
3.三羧酸循环是单向反应体系。
三羧酸循环中的柠檬酸合成酶、异柠檬酸脱氢酶、α-酮戊二酸脱氢酶复合体是该代谢途径的限速酶,所催化的是单向不可逆反应,所以三羧酸循环是不能逆转的,这保证了线粒体供能系统的稳定性。
4.三羧酸循环必须不断补充中间产物。
三羧酸循环的中间产物,从理论上讲,可以循环不消耗,但是由于体内各代谢途径的相互交汇和转化,三羧酸循环的中间产物常移出循环而参与其它代谢途径,如草酰乙酸可转变为天冬氨酸而参与蛋白质合成,琥珀酰辅酶A可用于血红素的合成,α-酮戊二酸可转变为谷氨酸等。
所以为了维持三羧酸循环中间产物的一定浓度,保证三羧酸循环的正常运转,就必须补充消耗的中间产物,称为回补反应。草酰乙酸的浓度,直接与乙酰辅酶A进入三羧酸循环的速度有关,因此不断补充草酰乙酸是使三羧酸循环得以顺利进行的关键,因而由丙酮酸形成草酰乙酸是最重要的回补反应。
草酰乙酸可以脱羧生成磷酸烯醇式丙酮酸,后者可以在肝脏和肾脏中逆糖酵解而生成葡萄糖或糖原,这就是非糖物质转化为糖的途径,称为【糖异生】,因此,三羧酸循环中所有的中间产物都能异生成糖。注意,乙酰CoA是无法异生成糖的,为什么?
5.三羧酸循环是三大营养物质彻底氧化的共同通路,是氧化释放能量产生ATP最多的阶段。糖、脂肪、蛋白质在体内氧化分解都将产生乙酰辅酶A,然后进入三羧酸循环进行降解,因此三羧酸循环是三大营养素在体内氧化的最终代谢通路(图5-1-1)。循环中脱下的四对氢,可进入呼吸链氧化磷酸化生成ATP,是体内ATP生成最多的反应阶段。
6.三羧酸循环是体内三大物质互变的枢纽
三羧酸循环是糖、脂肪、氨基酸代谢联系的枢纽。如葡萄糖氧化分解可生成乙酰CoA和NADPH+H,而乙酰CoA和NADPH+H可合成脂肪酸,进而合成脂肪;糖和甘油在体内代谢可生成α-酮戊二酸及草酰乙酸等三羧酸循环的中间产物,这些中间产物可以转变成为某些氨基酸;而有些氨基酸又可通过脱氨基作用生成α-酮酸,再经糖异生的途径生成糖或转变成甘油,因此三羧酸循环不仅是三大营养物质分解代谢的最终共同途径,而且也是它们互变的枢纽。
氢的燃烧三大营养物质在体内的氧化可以分为三个阶段,首先是糖、脂肪、蛋白质经过分解代谢生成乙酰辅酶A;接着乙酰辅酶A进入三羧酸循环脱羧脱氢,生成CO2并使NAD和FAD还原成NADH+H和FADH2;第三阶段是NADH+H和FADH2中的氢经呼吸链传给氧生成水,氧化过程中释放出来的能量合成ATP。前两个阶段已在前讲述,本节主要讨论第三阶段,即代谢物脱下的氢是如何通过呼吸链传递给氧生成水,细胞通过什么方式将氧化过程中释放的能量转变成ATP分子中的高能键。
(一)呼吸链(电子传递链)
代谢物脱下的氢通过多种酶和辅酶所催化的连锁反应逐步传递,最终与氧结合生成水,由于此过程与细胞利用O2生成CO2的呼吸有关,所以将此传递链称为呼吸链(respiratory chain)。在呼吸链中,酶和辅酶按一定的顺序排列在线粒体内膜上,其中传递氢的称为递氢体,传递电子的称为递电子体。
呼吸链由线粒体内膜上的5种复合体(复合蛋白)组成,它们是复合体I(NADH-Q还原酶,又称NADH氧化酶,辅基为FMN和Fe-S)、复合体II(琥珀酸-Q还原酶,辅基为FAD和Fe-S)、复合体III(细胞色素还原酶,辅基为血红素b、血红素c1和Fe-S)、复合体IV(细胞色素氧化酶,辅基血红素a、血红素a3和Cu)、复合体V(ATP合酶)。辅基传递氢和电子的有NAD、FMN、FAD、CoQ(图5-2-6),传递电子的有Fe-S(图5-2-7)和血红素Fe、Cu(图5-2-8),Fe、Cu通过得失电子来传递电子。
复合体I(NADH-Q还原酶)、CoQ、复合体III(细胞色素还原酶)和复合体IV(细胞色素氧化酶)组成NADH氧化呼吸链,复合体II(琥珀酸-Q还原酶)、复合体III(细胞色素还原酶)和复合体IV(细胞色素氧化酶)组成琥珀酸氧化呼吸链。
(二)电子传递与线粒体内膜两侧质子梯度的形成
【NADH氧化呼吸链的电子(氢)传递】线粒体内大多数脱氢酶都以NAD作为辅酶,在脱氢酶催化下底物SH2脱下的氢交给NAD生成NADH+H。NADH在NADH-Q还原酶(复合体I)作用下,NADH+H将氢原子传递给FMN生成FMNH2,后者再将氢传递给Q生成QH2,此时两个氢原子解离成2个质子和2个电子,2个质子游离于介质中,2个电子经由细胞色素还原酶(复合体III)传递至细胞色素C,然后细胞色素氧化酶(复合体IV)将细胞色素C上的2个电子传递给氧生成O,O与2H结合生成水。
【琥珀酸氧化呼吸链的电子(氢)传递】琥珀酸-Q还原酶使琥珀酸脱氢生成FADH2,然后将FADH2上的氢传递给Q生成QH2,其后的传递过程如NADH呼吸链。
电子传递的氧化势能使线粒体基质的氢离子H泵出到膜间腔(内膜外侧),从而形成内膜两侧的质子梯度差(内正外负),这个梯度差是合成ATP的势能所在。
复合体I(NADH-Q还原酶)、复合体III(细胞色素还原酶)、复合体IV(细胞色素氧化酶)具有质子泵作用,复合体II(琥珀酸-Q还原酶)传递电子的氧化势能太小,不能将质子泵出。
(三)ATP的合成
ATP合酶(复合体V)由FO和F1等蛋白复合而成,有质子通道,当质子由膜间腔经质子通道回流时,ATP合酶被活化而驱动ATP的合成(图5-2-11)。这种合成ATP的方式称做【氧化磷酸化】,即在呼吸链中伴随电子(质子)传递的氧化还原反应产生的电化学势能,推动ADP磷酸化为ATP的过程。氧化磷酸化是细胞合成ATP的主要方式,合成ATP的另一种方式是【底物磷酸化】(见本章糖酵解一节)。注意:ATP合酶的FO中的O是字母O,不是零。
新近的研究表明,NADH经呼吸链氧化可合成2.5个ATP分子,FADH2经呼吸链氧化可生成1.5个ATP分子。注意,这与传统的数字是不同的!
ATP是人体各种生命活动能量的直接供给者,它是食物中蕴藏的能量和机体利用的能量之间的纽带。糖、脂类及蛋白质等能源物质氧化分解释放的能量约有40%以化学能贮存在高能化合物中,ATP是体内最重要的高能化合物。但是ATP在细胞中的含量很低,在哺乳动物的脑和肌肉中约为3~8mmol/kg,这么微的含量只能供给肌肉剧烈活动1s左右的消耗,因此肌肉和脑中存在着一类贮存能量的高能化合物,这个化合物就是磷酸肌酸。
ATP将~P转移给肌酸(CP)生成磷酸肌酸(CP~P),作为肌肉和脑组织中能量的一种贮存形式。磷酸肌酸在脑中的含量是ATP的1.5倍,肌肉中相当于ATP的5倍。受过良好训练的运动员肌肉中的磷酸肌酸含量可高达30mmol/kg。当机体消耗ATP过多时,磷酸肌酸将~P转移给ADP,生成ATP,供生理活动之用。磷酸肌酸是细胞内首先供应ADP使之再合成ATP的能源物质。ATP是体内的能量使者,是生物体内能量的储存和利用的中心分子。
(四)氧化磷酸化的影响因素?
1.ADP/ATP比值的影响
氧化磷酸化主要受细胞对能量需求的影响。细胞能量供应缺乏时,即ATP减少,ADP增加,ADP/ATP比值增大,氧化磷酸化速率加快,NADH迅速减少而NAD增多,促进三羧酸循环;反之,细胞内能量供应充足时,即ATP增加,ADP减少,ADP/ATP比值减少,氧化磷酸化速率减慢,NADH消耗减少,三羧酸循环减缓。ADP/ATP比值是调节氧化磷酸化的基本因素,这种反馈调节可使机体适应生理需要,合理利用能源。
2.甲状腺素的调节
甲状腺素能诱导细胞膜上Na-K-ATP酶的生成,使ATP分解为ADP的速度加快,线粒体中ADP/ATP比值增大,导致氧化磷酸化加强。由于ATP的合成和分解都增加,使机体耗氧量和产热量都增加。所以甲状腺功能亢进患者常出现基础代谢率增高。
3.抑制剂的作用
一些化合物对氧化磷酸化有抑制作用,根据其作用部位不同分为两类:
⑴呼吸链抑制剂:
这类抑制剂阻断呼吸链上某一环节的电子传递。由于电子传递受阻,磷酸化反应也无法正常进行。如安眠药异戊巴比妥(阿米妥)、植物毒素鱼藤酮(常用作杀虫剂)能阻断NADH到Q之间的电子传递;一氧化碳和氰化物能与细胞色素氧化酶分子中的铁离子结合,使其失去电子传递能力。
⑵解偶联剂:
这类抑制剂不影响呼吸链的电子传递,而是解除氧化与磷酸化的偶联作用,使氧化过程产生的能量不能生成ATP,而是以热能的形式散发。如2,4-二硝基酚、双香豆素等属于解偶联剂,感冒或患某些传染病时体温升高,是由于细菌或病毒产生的解偶联剂所致。
NADH的氧化体内很多物质氧化分解产生NADH,线粒体内生成的NADH可直接通过呼吸链进行氧化磷酸化,而胞液中生成的NADH由于不能自由透过线粒体内膜,故需通过某种转运机制,将氢转移到线粒体内,重新生成NADH或FADH2后再参加氧化磷酸化。这种转运机制主要有α-磷酸甘油穿梭和苹果酸穿梭。
(一)3-磷酸甘油(α-磷酸甘油)穿梭系统
该穿梭系统主要存在于肌肉和神经组织,它是通过α-磷酸甘油将胞液中NADH的氢带入线粒体内,具体过程如下:
当胞液中NADH浓度升高时,磷酸二羟丙酮在胞液α-磷酸甘油脱氢酶(辅酶为NAD)催化下由NADH+H供氢生成α-磷酸甘油,后者进入线粒体后在线粒体内α-磷酸甘油脱氢酶(辅酶为FAD)的催化下重新生成磷酸二羟丙酮和FADH2。磷酸二羟丙酮穿出线粒体外可继续利用。生成的FADH2经呼吸链氧化磷酸化,这种穿梭作用可生成1.5分子ATP。
(二)苹果酸穿梭系统
又称苹果酸-天冬氨酸穿梭系统,主要存在于肝、肾、心。是指通过苹果酸将胞液中NADH的氢带进线粒体内,具体过程如下:
当胞液中NADH浓度升高时,首先由苹果酸脱氢酶(辅酶NAD)催化,使草酰乙酸还原成苹果酸。苹果酸在线粒体内膜转位酶的催化下穿过线粒体内膜,进入线粒体的苹果酸,在苹果酸脱氢酶作用下脱氢生成草酰乙酸,并生成NADH+H。生成的NADH+H通过呼吸链传递进行氧化磷酸化,生成2.5分子ATP。
草酰乙酸不能直接透过线粒体内膜返回胞液,但它可在天冬氨酸转氨酶作用下从谷氨酸接受氨基生成天冬氨酸,谷氨酸转出氨基后生成α酮戊二酸,α酮戊二酸、天冬氨酸都能在膜上转位酶的作用下穿过线粒体内膜而进入胞液,在胞液中天冬氨酸和α酮戊二酸在天冬氨酸转氨酶的作用下又重新生成草酰乙酸和谷氨酸,草酰乙酸又可重新参与苹果酸穿梭作用。
ATP的总结算(一)葡萄糖彻底氧化产生ATP结算
关于葡萄糖彻底氧化为水和二氧化碳究竟产生多少ATP分子的问题一直受到人们的关注。葡萄糖分解通过糖酵解和柠檬酸循环的底物磷酸化作用产生ATP的分子数,根据化学计算可以得到明确的答复。但是氧化磷酸化产生的ATP分子数并不十分准确。因为质子泵、ATP合成以及代谢物的转运过程并不需要是完整的数值甚至不需要固定值。根据最新测定计算,一对NADH传至O2,所产生的ATP分子数是2.5个,琥珀酸及脂肪酸氧化产生的FADH2传递至O2,产生的ATP是1.5个。这样,当一分子葡萄糖彻底氧化彻底氧化为CO2和H2O所得到的ATP分子数和过去传统的统计数(36或38个ATP)少了6个ATP分子,成为30个或32个。全部的统计列于表5-2-1。在30或32个ATP分子中,氧化磷酸化产生26或28个,底物磷酸化产生6个,葡萄糖活化共消耗2个。
(二)软脂酸彻底氧化产生ATP结算
软脂酸(16碳饱和脂肪酸)彻底氧化产生ATP数目为106个,其中氧化磷酸化为100个,底物磷酸化为8个,脂肪酸活化消耗2个(消耗1个ATP生成1个AMP,即消耗2个高能键)。
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国