

变态反应也叫超敏反应。是指免疫系统对一些对机体无危害性的物质如花粉、动物皮毛等过于敏感,发生免疫应答,对机体造成伤害。人们日常遇到的皮肤过敏,皮肤骚痒、红肿,就是一种变态反应。
概述若机体已被某种寄生虫抗原致敏,当再次接触相同抗原时则二次免疫应答增强,或长期受染,早期过去后的机体反应相似于二次免疫应答反应。因免疫应答过强而导致组织损伤(免疫病理变化),即称为变态反应(allergy),或超敏反应(hypersensitivityreaction)。1963年起Gell与Coombs按变态反应发生发展的近代知识,首先提出四型分型法,即Ⅰ型——速发型(immediat type),Ⅱ型——细胞毒型(cytotoxic type)/细胞溶解型,Ⅲ型——免疫复合物型(immunecomplex type),以上3型均由抗体所介导;而Ⅳ型——迟发型(delayedtype)或细胞介导型(cellmediated type),由1所介导。
发生条件变态反应2的发生需要具备两个主要条件:
一是容易发生变态反应的特应性体质,这是先天遗传决定的,并可传给下代,其机率遵循遗传法则;
二是与抗原的接触,有特应性体质的人与抗原首次接触时即可被致敏,但不产生临床反应,被致敏的机体再次接触同一抗原时,就可发生反应,其时间不定,快者可在再次接触后数秒钟内发生,慢者需数天甚至数月的时间。
特点和常见病变态反应的是伴有炎症反应和组织损伤。
2005年,在世界首个过敏性疾病日,世界变态反应组织公布对30个国家进行流行病学调查结果显示:在这些国家的12亿总人口中,22%(2亿5 千万人)患有免疫球蛋白E(IgE)介导的过敏性疾病,如过敏性鼻炎、哮喘、结膜炎、湿疹、食物过敏、药物过敏和严重过敏反应等。
流行病学在西方国家,过敏性鼻炎和哮喘的患病率近40年内迅速增加,欧洲普通人群过敏性疾病的流行病调查开始于上世纪初,其患病率在20 年代低于1%,工业革命以后从50年代至80年代逐渐上升,80年代以后戏剧性上升。瑞士过敏性鼻炎的发病率1926年为0.82%,1958年为 4.8%,1985年9.6%,1995年跃升为14.2%。美国一项依据皮肤试验的调查显示:大约4000~5000万人有过敏问题,其中3950万人患有季节性过敏性鼻炎。据世界卫生组织(WHO)估计,全球约有1亿5千万人患有哮喘,其中50%以上的成人及至少80%的儿童患者均由过敏因素诱发,每年有180000多人死于哮喘。
食物过敏、湿疹和药物过敏的患病率近年也明显升高,美国有30 0万人对花生和坚果过敏,6岁以下儿童食物过敏的患病率为4%,成人为1%~2%。食物过敏已成为严重过敏反应和过敏性休克的主要原因。特应性皮炎在欧美国家和亚太发达地区的婴幼儿和儿童中很常见,其发病率已从60年代的3%上升为90年代的10%。急性荨麻疹影响10 %~20%美国人的生活,其中50%的症状持续6个月以上。美国药物过敏占全部药物不良反应的10%,青霉素是最常见的致敏原因,每年有400人因青霉素过敏性休克而死亡。
过敏性疾病患病率的迅速增高,已达到某种流行病的程度。这与长期的、持续的环境因素和生活方式的改变有关。西方学者发现,过敏性疾病的发病在发达国家和地区高于发展中国家,城市高于乡村,污染地区高于非污染地区;在发展中国家,则与采用城市化的“西方” 生活方式相关,农民的孩子较其他孩子较少患过敏性疾病;在城市,父母是高薪阶层或专业人士的子女较低薪阶层的子女更容易罹患过敏性疾病。东西德统一之前,西德过敏性疾病的发病率明显高于东德,但在统一多年后的今天,过敏性疾病的发病情况已没有区别。
分类Ⅰ型变态反应即速发型(Ⅰ型变态反应),又称过敏反应,是临床最常见的一种,其特点是:由IgE介导,肥大细胞和嗜碱粒细胞等效应细胞以释放生物活性介质的方式参与反应;发生快,消退亦快;常表现为生理功能紊乱,而无严重的组织损伤;有明显的个体差异和遗传倾向。
过敏原进入机体后,诱导B细胞产生IgE抗体。IgE与靶细胞有高度的亲和力,牢固地吸附在肥大细胞、嗜碱粒细胞表面。当相同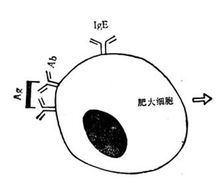 的抗原再次进入致敏的机体,与IgE抗体结合,就会引发细胞膜的一系列生物化学反应,启动两个平行发生的过程:脱颗粒与合成新的介质。①肥大细胞与嗜碱粒细胞产生脱颗粒变化,从颗粒中释放出许多活性介质,如组胺、蛋白水解酶、肝素、趋化因子等;②同时细胞膜磷脂降解,释放出花生四烯酸。它以两条途径代谢,分别合成前列腺素、血栓素A2;和白细胞三烯(LTs)、血小板活化因子(PAF)。各种介质随血流散布至全身,作用于皮肤、粘膜、呼吸道等效应器官,引起小血管及毛细血管扩张,毛细血管通透性增加,平滑肌收缩,腺体分泌增加,嗜酸粒细胞增多、浸润,可引起皮肤粘膜过敏症(荨麻疹、湿疹、血管神经性水肿),呼吸道过敏反应(过敏性鼻炎、支气管哮喘、喉头水肿),消化道过敏症(食物过敏性胃肠炎),全身过敏症(过敏性休克),小结:由于IgE多由粘膜分泌,所以I型多引起粘膜反应。
的抗原再次进入致敏的机体,与IgE抗体结合,就会引发细胞膜的一系列生物化学反应,启动两个平行发生的过程:脱颗粒与合成新的介质。①肥大细胞与嗜碱粒细胞产生脱颗粒变化,从颗粒中释放出许多活性介质,如组胺、蛋白水解酶、肝素、趋化因子等;②同时细胞膜磷脂降解,释放出花生四烯酸。它以两条途径代谢,分别合成前列腺素、血栓素A2;和白细胞三烯(LTs)、血小板活化因子(PAF)。各种介质随血流散布至全身,作用于皮肤、粘膜、呼吸道等效应器官,引起小血管及毛细血管扩张,毛细血管通透性增加,平滑肌收缩,腺体分泌增加,嗜酸粒细胞增多、浸润,可引起皮肤粘膜过敏症(荨麻疹、湿疹、血管神经性水肿),呼吸道过敏反应(过敏性鼻炎、支气管哮喘、喉头水肿),消化道过敏症(食物过敏性胃肠炎),全身过敏症(过敏性休克),小结:由于IgE多由粘膜分泌,所以I型多引起粘膜反应。
Ⅱ型变态反应即细胞毒型(Ⅱ型变态反应),抗体(多属IgG、少数为IgM、IgA)首先同细胞本身抗原成分或吸附于膜表面成分相结合,然后通过四种不同的途径杀伤靶细胞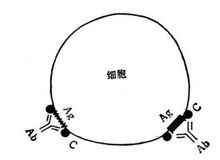 。
。
(1)抗体和补体介导的细胞溶解:IgG/IgM类抗体同靶细胞上的抗原特异性结合后,经过经典途径激活补体系统,最后形成膜攻击单位,引起膜损伤,从而靶细胞溶解死亡。
(2)炎症细胞的募集和活化:补体活化产生的过敏毒素C3a、C5a对中性粒细胞和单核细胞具有趋化作用。这两类细胞的表面有IgG Fc受体,故IgG与之结合并激活它们,活化的中性粒细胞和单核细胞产生水解酶和细胞因子等从而引起细胞或组织损伤。
(3)免疫调理作用:与靶细胞表面抗原结合的IgG抗体Fc片段同巨噬细胞表面的Fc受体结合,以及C3b促进巨噬细胞对靶细胞的吞噬作用。
(4)抗体依赖细胞介导的细胞毒作用:靶细胞表面所结合的抗体的Fc段与NK细胞、中性粒细胞、单核—巨噬细胞上的Fc受体结合,使它们活化,发挥细胞外非吞噬杀伤作用,使靶细胞破坏。
Ⅲ型变态反应即免疫复合物型(Ⅲ型变态反应),又称血管炎型超敏反应。其主要特点是:游离抗原与相应抗体结合形成免疫复合物(IC),若IC不能被及时清除,即可在局部沉积,通过激活补体,并在血小板、中性粒细胞及其他细胞参与下,引发一系列连锁反应而致组织损伤。
在免疫应答过程中,抗原抗体复合物的形成是一种常见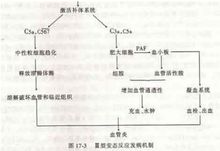 现象,但大多数可被机体的免疫系统清除。如果因为某些因素造成大量复合物沉积在组织中,则引起组织损伤和出现相关的免疫复合物病,中性粒细胞聚集释放溶酶体酶是造成组织损伤的直接原因。
现象,但大多数可被机体的免疫系统清除。如果因为某些因素造成大量复合物沉积在组织中,则引起组织损伤和出现相关的免疫复合物病,中性粒细胞聚集释放溶酶体酶是造成组织损伤的直接原因。
免疫复合物沉积的影响因素有如下几个:
(1)循环免疫复合物的大小:这是一个主要因素,一般来讲分子量为约1000kD沉降系数为8.5—19S的中等大小的可溶性免疫复合物易沉积在组织中。
(2)机体清除免疫复合物的能力:它同免疫复合物在组织中的沉积程度呈反比。
(3)抗原和抗体的理化性质:复合物中的抗原如带正电荷,那么这种复合物就很容易与肾小球基底膜上带负电荷的成分相结合,因而沉积在基底膜上。
(4)解剖和血流动力学因素:对于决定复合物的沉积位置是重要的。肾小球和滑膜中的毛细血管是在高流体静压下通过毛细血管壁而超过滤的,因此它们成为复合物最常沉积的部位之一。
(5)炎症介质的作用:活性介质使血管通透性增加,增加了复合物在血管壁的沉积。
(6)抗原抗体的相对比例:抗体过剩或轻度抗原过剩的复合物迅速沉积在抗原进入的局部。
常见的Ⅲ型变态反应疾病有:Arthus反应、一次血清病、链球菌感染后肾小球肾炎等。
Ⅳ型变态反应即迟发型(Ⅳ型变态反应),与上述由特异性抗体介导的三型变态反应不同,Ⅳ型是由特异性致敏效应T细胞介导的。此型反应局部炎症变化出现缓慢,接触抗原24—48h后才出现高峰反应,故称迟发型变态反应。机体初次接触抗原后,T细胞转化为致敏淋巴细胞,使机体处于过敏状态。当相同抗原再次进入时,致敏T细胞识别抗原,出现分化、增殖,并释放出许多淋巴因子,吸引、聚集并形成以单核细胞浸润为主的炎症反应,甚至引起组织坏死。 常见Ⅳ型变态反应有:接触性皮炎、移植排斥反应、多种细菌、病毒(如结核杆菌、麻 疹病毒)感染过程中出现的Ⅳ型变态反应等。
疹病毒)感染过程中出现的Ⅳ型变态反应等。
在寄生虫方面,利什曼原虫引起的皮肤结节,有明显的细胞反应和肉芽肿形成。血吸虫排出的虫卵随血液流入肝脏,毛蚴成熟分泌可溶性抗原,经卵壳微孔释出,使淋巴细胞致敏,当再接触抗原时,致敏的T淋巴细胞放出淋巴毒素(LT),巨噬细胞移动抑制因子((MIF),嗜酸粒细胞趋化因子(ECF-A),因此在虫卵周围出现以淋巴细胞、巨噬细胞、嗜酸性粒细胞浸润为主的肉芽肿。在寄生虫感染中,有的寄生虫病可同时存在多型变态反应,病理后果是多种免疫病理机制的复合效应,甚为复杂多变。如前已述及,血吸虫感染时引起的尾蚴性皮炎(属I型和IV型变态反应)、对童虫杀伤的ADCC作用(属Ⅱ型变态反应)、血吸虫性肾小球肾炎(属Ⅲ型变态反应)以及血吸虫虫卵性肉芽肿(属IV型变态反应)。又如由昆虫引起的变态反应,主要是速发型和迟发型,部分是免疫复合物型。当昆虫叮咬时它的分泌物、排泄物以及毒毛等(过敏原)侵入人体而诱发局部的和全身的变态反应现象。
鉴别疥螨疥螨在进食的吸吮过程中,向皮肤内分泌唾液,其唾液中或者用来叮咬的口器中含有的抗原诱发免疫应答。
Ⅰ型变态反应:疥螨在皮肤上掘隧道时抗原即被导入,这些抗原通过表皮向下扩散;针对疥螨特异性抗原的IgE循环,渗入真皮,其Fc段接触肥大细胞的Fc受体;当疥螨抗原同肥大细胞表面的特异性IgE相遇并互相作用,导致脱颗粒,引起血管扩张和血浆漏出。
IgE或I型反应可能参与疥疮的发病机制,其证据有:
①患者的IgE水平升高;
②经有效的治疗后IgE即下降;
③皮内注射成熟雌疥螨浸出物可使以前患过疥疮者发生风团反应,而健康志愿者则否;
④该反应可通过一种血清因子而被动转移;
⑤结痂性疥疮血清在体外可致组胺的释放和嗜碱粒细胞的脱颗粒;
⑥结痂性疥疮血清IgE明显升高;
⑦用粗制疥螨抗原进行放射免疫吸附试验显示血清中抗原特异性IgE升高;
⑧针对疥螨的IgE抗体能与一种普通的屋尘螨(dermatophagoidespteronssinus)的抗原发生交叉反应。
Ⅱ型变态反应:由IgG或IgM结合抗原而产生。常由补体及吞噬细胞介导。当补体存在时,正常人血清能否直接杀伤疥螨,尚无关键性研究来证实。某些疥疮患者血清IgG与IgM升高,但与皮损范围、严重程度或病期并不相关。然而,在有效的治疗后可下降。这些高水平的IgG和IgM是抗原特异性的,还是非特异性的,抑或是否与伴发的脓皮性细菌感染有关,还不清楚。有时患者血清IgA下降,治疗后即升高,其降低的原因不明。
补体C3、C4一般正常,但并不意味着补体不被激活。直接免疫荧光检查发现患部皮肤的表皮真皮连接处有IgM和C3沉积,提示来自疥螨的抗原向下弥散,来自真皮血管的抗体向上弥散,二者发生反应。然而,IgM和C3的沉积也可见于其它许多不同原因的炎症性皮肤病。因此,关于疥疮表皮真皮连接处免疫反应物的意义仅属推测性的,沉积有可能继发于炎症。
Ⅲ型变态反应:在预先致敏的宿主,皮内注射抗原可以实验性诱发Arthus反应。疥疮患者可能有针对疥螨各种抗原的IgG或IgM抗体。如通过搔抓或其它机制抗原进入真皮,而同时宿主有适量IgG和IgM抗体的话,那么在理论上则可能发生局部Arthus反应。循环免疫复合物沉积于皮肤小静脉也能引起血管炎。这些复合物激活补体导致吞噬细胞浸润血管壁,从而引起血管坏死。
在治疗前后的结痂性疥疮与普通疥疮患者血清中已发现由疥螨抗原及抗体组成的免疫复合物。复合物中抗原的性质尚不明确,疥疮感染可引起全身性皮肤血管炎,活检标本已发现血管炎伴有血管壁坏死。疥疮的丘疹、结节和隧道处的血管壁可有C3、IgM及IgA沉积。在成熟或未成熟疥螨附近可发生表皮坏死。虽然丘疹发生的溃烂多数是由于搔抓所引起的,但有些可能是由于局部Arthus反应及伴发的坏死性炎症所致。
Ⅳ型变态反应:Mellanby将疥螨接种于健康的志愿者。第一个月,受试者无症状,搔痒性 丘疹往往在初次接种后一个月或更长时间才出现;第二次接种时,症状及体征迅速出现,没有初次接种那样长的潜伏时间,这提示疥疮的症状及体征是免疫应答导致的,属典型的由T淋巴细胞介导的迟发性超敏反应。疥疮是由人疥螨感染皮肤导致的,在缺乏免疫反应时,疥螨本身导致的症状极少。当机体对其产生免疫应答时,患者出现丘疹、风团、结节及结痂性丘疹,这些损害是机体对疥螨抗原发生迟发性超敏反应的结果。红斑、风团及极少发生的血管炎可能是由IgE或(和)IgM及IgG参与的体液免疫所致。
丘疹往往在初次接种后一个月或更长时间才出现;第二次接种时,症状及体征迅速出现,没有初次接种那样长的潜伏时间,这提示疥疮的症状及体征是免疫应答导致的,属典型的由T淋巴细胞介导的迟发性超敏反应。疥疮是由人疥螨感染皮肤导致的,在缺乏免疫反应时,疥螨本身导致的症状极少。当机体对其产生免疫应答时,患者出现丘疹、风团、结节及结痂性丘疹,这些损害是机体对疥螨抗原发生迟发性超敏反应的结果。红斑、风团及极少发生的血管炎可能是由IgE或(和)IgM及IgG参与的体液免疫所致。
肠阿米巴病溶组织内阿米巴(Entamoebahistolytica)(以下简称阿米巴)它可引起肠阿米巴病或肠外阿米巴病(主要是阿米巴肝脓肿)。在阿米巴肝脓肿(amoebicliverabscess,ALA))时细胞免疫抗阿米巴的机制简述如下。有人指出,细胞免疫在肝肠之间构成一个免疫学屏障(immunologicalbarrier),如果这种屏障破坏,则可导致ALA的发生。那么这种屏障是如何发挥作用的呢?很多资料表明巨噬细胞、淋巴细胞在这种屏障中发挥着重要的作用。Chugh(1985)和Saxena(1966)观察到单个核细胞(mononuclearcell,MNC)及腹腔巨噬细胞与滋养体之间有直接接触。效应细胞先是连接到滋养体上,然后又从死亡的滋养体游走开来,证明杀阿米巴滋养体的为ADCC,且一个效应细胞能杀伤多个靶细胞。ADCC不需要补体的参与,但需要低浓度的抗体共同作用。抗体通过Fc段连接到效应细胞上,再经Fab段与靶细胞连接,这样可把大量的效应细胞连接到滋养体上,通常是完全包绕滋养体,从而使滋养体变形、解体,然后,效应细胞又可在抗体参与下杀伤其它滋养体。Chadee等(1985)用感染阿米巴的沙鼠的腹腔巨噬细胞及淋巴细胞(来源于肠系膜淋巴结)进行体外研究,也证明效应细胞对滋养体的杀伤系ADCC。参与阿米巴滋养体的ADCC的效应细胞,文献报告最多的是巨噬细胞,其次是淋巴细胞、细胞毒T细胞和K细胞,而Chugh认为是单核细胞。参与的抗体为IgG。Saxena用2—巯基乙醇处理脉鼠抗阿米巴血清(破坏IgM),对效应细胞抗阿米巴的能力无影响。纯化的IgM不能引起ADCC,故认为参与的抗体为IgG。同时,在实验中,经SephadexG—200过柱的阿米巴抗原中,用其组分I免疫动物(MW650000)所产生的免疫血清,IgG滴度最高。
隐孢子虫感染人类引起隐孢于虫病的病原为微小隐孢子虫(Cryptosporidiumparvum)。隐孢子虫抗原主要分卵囊壁抗原和子孢子抗原,目前隐孢子虫病的免疫机制尚不清楚。多数学者认为以细胞免疫为主。不少资料表明,凡是细胞免疫功能正常的隐孢子虫病人一般都表现为急性自限性腹泻,而细胞免疫功能受损者则为慢性持续性腹泻。摘除胸腺而缺乏T淋巴细胞的小鼠,经实验感染隐孢子虫后不能消除卵囊,而胸腺机能正常的小鼠则表现为一过性感染。Moon发现乳源性抗隐孢子虫IgG、IgM不能保护小鼠抵抗隐孢子虫感染,因而被认为隐孢子虫病的免疫主要依赖细胞而非抗体。但也有持不同意见者,Chrisp用豚鼠作动物模型,发现成鼠、幼鼠口服接种隐抱子虫卵囊后同样易感,并产生特异性体液免疫,已产生体液免疫的豚鼠再接种隐孢子虫卵囊,不发生再感染。Riggs认为免疫血清对识别及中和子抱子抗原具有一定作用。Laxer通过检测儿童血清、粪便及十二指肠液中的特异性隐孢子虫IgA、IgM、IgG抗体,并以迟发型皮肤过敏反应检测细胞免疫功能,得出的结论为隐孢子虫病的免疫机制可能为抗体依赖的细胞介导的细胞毒作用(ADCC)。
分型免疫成分损伤机制寄生虫感染举例ⅠIgE肥大细胞,嗜碱性粒细胞及其介质血吸虫尾蚴蚴性皮炎,热带肺嗜酸性粒细胞增多症,包虫囊破裂所致的休克。
ⅡIgM,IgG补体活化,白细胞趋化、活化,NK细胞作用疟疾(三日疟)的贫血,恰加斯病心肌炎。
ⅢCAg补体活化,白细胞趋化、活化疟疾(三日疟)肾病综合征,急性血吸虫病。
IVCD8+T细胞CD4+T细胞直接致靶细胞溶解,活化吞噬细胞、细胞因子释放导致炎症皮肤利什曼病,血吸虫尾蚴性皮炎、肝硬化,丝虫性象皮肿。
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国