

乙酰胆,是一种神经递质。在组织内迅速被胆碱酯酶破坏。乙酰胆碱能特异性地作用于各类胆碱受体,但其作用广泛,选择性不高。临床不作为药用,一般只做实验用药。在神经细胞中,乙酰胆碱是由胆碱和乙酰辅酶A在胆碱乙酰移位酶(胆碱乙酰化酶)的催化作用下合成的。主流研究认为人体内该物质含量增多与阿尔兹海默病(老年痴呆症)的症状改善显著相关。
简介定义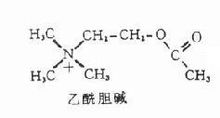
乙酰胆碱,分子式CH3COOCH2CH2N+(CH3)3在神经细胞中,乙酰胆碱是由胆碱和乙酰辅酶A在胆碱乙酰移位酶(胆碱乙酰化酶)的催化作用下合成的。由于该酶存在于胞浆中,因此乙酰胆碱在胞浆中合成,合成后由小泡摄取并贮存起来。
进入突触间隙的乙酰胆碱作用于突触后膜发挥生理作用后(乙酰胆碱可引起受体膜产生动作电位),就被胆碱酯酶水解成胆碱和乙酸,这样乙酰胆碱就被破坏而推动了作用(迅速分解是为了避免受体细胞膜持续去极化而造成的传导阻滞),这一过程称为失活。
引起乙酰胆碱量子性释放的关键因素是神经末梢去极化引起的Ca2+内流。当神经冲动传至神经终板时,膜电位下降,导致可使Ca2+通过的电压闸门通道开放, 使Ca2+进入终板,从而刺激终板分泌乙酰胆碱。乙酰胆碱再进一步作用于肌细胞导致肌细胞收缩。
人类对乙酰胆碱的认识1914年,Ewins在麦角菌中发现了乙酰胆碱,这是首次在非神经细胞中发现乙酰胆碱的报道。随后,人们陆续在多种细菌、真菌、低等植物和高等植物中发现了乙酰胆碱及其相关的酶和受体。随着胆碱能系统在植物中的发现和研究的深入,人们似乎有望在分子水平发现动植物间的又一相似性,因而植物学家抱着极大的热情投入了这方面的研究。但是由于当时研究手段的限制、对动植物之间的差别认识不足,以及某些研究在其它的实验室难以重复的缘故,使得植物乙酰胆碱的研究多处于零星的、非系统的状态,研究的深度和广度远远无法与动物相比。尚未对其在植物中的作用机理提出一个合理的解释。 近年来,我们和国外其它几家实验室重新开展了乙酰胆碱在植物体内的生理作用和作用机理的研究,为揭示植物乙酰胆碱的作用机理提供了新的线索。1
作用植物生理过程种子萌发
乙酰胆碱和乙酰胆碱酯酶可能参与调控某些植物的种子萌发和幼苗早期生长,乙酰胆碱影响这些生理过程的机理可能涉及调控储藏物从下胚轴向植物快速生长部位的调运。乙酰胆碱对需光种子萌发的影响的研究有许多矛盾的报道。Tretyn等在研究乙酰胆碱及其类似物、乙酰胆碱酯酶抑制剂对不同光周期植物种子萌发的影响中发现,无论在光下还是在黑暗中这些化合物对光不敏感植物的种子萌发都没有影响。但在光下可以促进需光种子萌发,而在暗中则没有作用。对于不需光种子,乙酰胆碱抑制其在光下的萌发,乙酰胆碱类似物胆碱对上述过程则无影响。由于乙酰胆碱及乙酰胆碱酯酶在种子中广泛存在,因而有理由推测乙酰胆碱参与调控种子的萌发, 调控的机理可能涉及光对种子中乙酰胆碱酯酶活性的抑制。
生长
乙酰胆碱对于生长的影响因实验条件的不同,植物种类或同一植物不同组织而异。乙酰胆碱可以模拟红光的作用抑制大豆侧根的发育,还可以引起小麦幼苗生长和干重的增加。在离体组织中,乙酰胆碱可以刺激燕麦胚芽鞘和黄瓜下胚轴的伸长以及绿豆下胚轴的生长,刺激蚕豆下胚轴的生长而抑制其上胚轴的生长。总之,乙酰胆碱对植物生理过程的影响与所利用的组织及实验条件密切相关,其最大效应在pH酸性区。
成花作用
乙酰胆碱可以模拟红光的作用,抑制远红光刺激的过氧化物酶活性升高,从而使菠菜能在非诱导条件下开花。乙酰胆碱可以抑制连续光照条件下(24 h光照/0 h黑暗)长日浮萍G1的成花和刺激非诱导的短日条件下短日浮萍Torr的成花。阿托品可以抑制连续光照下生长的青萍G3成花而管箭毒则无作用,说明乙酰胆碱对成花的诱导作用可能是通过质膜上的类毒蕈碱型受体介导的。乙酰胆碱对成花的诱导作用还可能与它调控的膜对离子的通透性有关。光周期诱导的成花作用涉及到叶片膜电势的改变,乙酰胆碱还可能通过影响膜电势而参与成花诱导。
呼吸作用
乙酰胆碱可引起根尖细胞耗氧速率的增加。Jaffe以游离的线粒体为材料得到的结果已证实了这一点。伴随着氧的消耗,组织中ATP的水平下降10倍,自由磷水平升高14倍。乙酰胆碱的这种作用可能是其使呼吸的电子传递链与氧化磷酸化解偶联所造成。根据这些实验结果,Jaffe提出了乙酰胆碱对大豆根尖细胞的作用模式,即当胞间乙酰胆碱浓度升高时,乙酰胆碱到达其作用的靶部位,随后是分泌质子,氧的消耗和ATP水解增加,而这些过程均与膜对阳离子通透性的增加相关连。
光合作用
乙酰胆碱可以在不影响电子传递的情况下使叶绿体中的ATP合成下降80%以上。另外,浓度低于0.1 mmol的乙酰胆碱可以刺激非环式光合磷酸化的进行,而浓度大于0.1 mmol时非环式光合磷酸化则受抑制。在这两种情况下,乙酰胆碱并不影响NADP+的还原。新斯的明(neostigmine)可以抑制ATP的合成,但不影响电子从水到细胞色素f或NADP+的传递。毒蕈碱和阿托品同样可以抑制NADP+的还原和非环式光合磷酸化。
除此以外,乙酰胆碱还可以影响离体叶绿体对氧的吸收,抑制光刺激的叶绿体膨胀;刺激钠离子和钾离子从叶绿体流出。因而,乙酰胆碱在叶绿体中可能调控叶绿体膜对离子的通透性及电子传递和ATP合成间的偶联。
棚田效应红光促使黄化的绿豆和大麦根尖吸附到带负电的玻璃杯内壁上,而远红光则使根尖脱离杯壁释放到溶液中。这种现象称为棚田效应(tanada effect)。 在黑暗中乙酰胆碱可以使离体的大豆根尖吸附到带负电的玻璃杯内壁上,并阻止远红光引起的根尖脱离杯壁,乙酰胆碱酯酶抑制剂毒扁豆碱(eserine)增加组织对乙酰胆碱的敏感性。这些说明内源乙酰胆碱可能在这一生理过程中有控制作用。
红光可提高组织中乙酰胆碱水平,其原因可能与红光促进Pfr的形成有关,而后者则与乙酰胆碱合成有关。组织中乙酰胆碱水平升高可以刺激质子从根细胞流出到溶液中,从而形成表面正电势,以致根尖被吸附到带负电的玻璃杯内壁上;远红光促使光敏素从远红光吸收型(Pfr)转变为红光吸收型(Pr),致使根尖从玻璃杯内壁释放到溶液中。但是也有实验指出乙酰胆碱在这一过程中仅相当于单价阳离子的作用。
叶片运动Jaffe提出乙酰胆碱可能调控含羞草叶片的运动。紫花大翼豆是一种常用的牧草,在强光照下其叶片可以下垂以避免高光强对叶片的直接伤害。据报道,强光下来源于热带的品种比来自温带品种的叶片下垂快,光强减弱后下垂状态恢复更快。测定此种植物叶褥组织中乙酰胆碱的结果表明,乙酰胆碱水平的变化与叶片的状态密切相关。来源于热带品种的含量和变化幅度更大,外施乙酰胆碱可以使其从下垂状态中恢复。进一步研究的结果显示,叶片中乙酰胆碱水平的变化是由乙酰胆碱酯酶控制的,而乙酰胆碱酯酶主要分布于维管束的周围,因而推测乙酰胆碱可能通过影响离子进出维管束,进而影响水分的进出最终实现叶片运动的调控。
通透性乙酰胆碱可以刺激质子从大豆根尖细胞流出,诱导菠菜叶片膜电势的变化,抑制蓝光诱导的大豆下胚轴弯钩膜电势的超极化及该组织对钾的吸收,这些过程都涉及乙酰胆碱对膜透性的调节。
除了影响上述过程外,乙酰胆碱还可以影响组织对钙离子的吸收。Tretyn发现乙酰胆碱可以刺激黄化燕麦胚芽鞘对钙离子的吸收。乙酰胆碱酯酶的抑制剂可以增加组织对乙酰胆碱的敏感性;钙通道的抑制剂可以抑制乙酰胆碱刺激的钙吸收的增加。这些结果表明乙酰胆碱参与调控植物的钙通道。
代谢的影响乙酰胆碱可以影响植物的膜脂代谢。如它可以抑制磷掺入到黄化大豆茎切段的磷脂分子中,但在有氧条件下主要抑制磷掺入磷脂酰乙醇胺和磷脂酰胆碱,而在无氧条件下乙酰胆碱主要抑制磷掺入磷脂酰肌醇。这些结果表明植物的磷脂与动物的磷脂间有相似性,乙酰胆碱可以同样影响植物的磷脂代谢。
参与相互作用参与植物与植物以及细胞与细胞之间的相互作用
在一个生态环境中,植物与植物之间以及植物与其他生物之间常常表现出相互作用的关系。这种相互作用可以是促进性的也可以是抑制性的,即表现为相生相克的关系。乙酰胆碱酯酶存在于根瘤菌感染大豆所形成的根瘤中,而且乙酰胆碱酯酶的最大活性与根瘤对氮的最大同化期相一致,推测乙酰胆碱及其酯酶在根瘤菌和寄主植物间的相互作用中起一定作用。乙酰胆碱酯酶还存在于地衣的叶状体中,而且主要分布于组成地衣的真菌和藻类两种生物的界面。在其粉芽(soredia)产生孢子过程中,乙酰胆碱酯酶活性增加,而且酶活性集中分布在接触区。乙酰胆碱及乙酰胆碱酯酶参与地衣这种生物间相互作用的机理可能是通过调控膜对离子的通透性,并介导环境中光对地衣生殖影响而实现的 乙酰胆碱及乙酰胆碱酯酶还可能参与花粉与柱头间的识别作用。在裂叶牵牛中乙酰胆碱酯酶主要分布在雌蕊柱头的表面,还存在于花粉粒和花粉管的尖端。乙酰胆碱的激活剂和拮抗剂以及乙酰胆碱酯酶的抑制剂均可以影响某些植物的花粉萌发和花粉管伸长。因此推测乙酰胆碱和乙酰胆碱酯酶可能参与植物柱头和花粉间的相互作用。
作用机理在动物细胞中乙酰胆碱与受体结合后,一方面直接影响膜对离子的通透性,另一方面通过各种第二信使影响各种生理过程的进行。在植物界中,虽然乙酰胆碱的受体还没有在生化上最后确定,但是一系列药理学的证据表明植物中确实存在着乙酰胆碱的受体。关于植物中乙酰胆碱与受体结合后的事件了解甚少。有关乙酰胆碱在植物细胞中作用机理的研究,除上面的零散研究外,只有Tretyn等在小麦原生质体膨胀和幼叶展开中的研究较为系统,并揭示了其中的某些细节。
信号转导原生质体膨胀 红光可以刺激黄化小麦叶肉细胞原生质体体积膨胀,这种刺激作用可为随后的远红光照射所逆转,说明这一反应是在光敏素控制下进行的。红光对原生质体体积膨胀的刺激作用要求介质中含有Ca2+。乙酰胆碱可以代替红光在黑暗中引起原生质体的膨胀。与红光引起的反应不同,乙酰胆碱不仅可以在含Ca2+的介质中引起原生质体的膨胀,而且在含Na+或K+的介质中也可以引起原生质体的膨胀。
除乙酰胆碱外,只有氨基甲酰胆碱可以刺激原生质体的膨胀,而胆碱、丙酰胆碱和丁酰胆碱则无此作用。乙酰胆碱酯酶的抑制剂毒扁豆碱可以增加原生质体对乙酰胆碱的敏感性。据此可以认为乙酰胆碱能特异地刺激黄化小麦叶肉原生质体膨胀
乙酰胆碱诱导原生质体膨胀过程中是否涉及乙酰胆碱受体的参与可用乙酰胆碱受体的激活剂和抑制剂来确定。乙酰胆碱N型受体的激活剂烟碱在含Na+或K+的介质中可以直接刺激原生质体膨胀,而在含Ca2+的介质中,烟碱没有作用。与以上结果不同,M型受体的激活剂毒蕈碱可以在含Ca2+的介质中刺激原生质体膨胀,而在含Na+或K+的介质中没有作用。采用乙酰胆碱受体抑制剂的研究也得出同样的结论。M型受体的抑制剂阿托品在含Na+或K+的介质中对乙酰胆碱刺激的原生质体膨胀没有作用,但在含Ca2+介质中则可以抑制乙酰胆碱诱导的原生质体膨胀。N型受体抑制剂管箭毒在含Ca2+介质中对乙酰胆碱刺激的原生质体膨胀没有作用,但在含Na+或K+的介质中则有抑制作用。荧光定位技术证明N型乙酰胆碱受体主要分布在原生质体表面。
在乙酰胆碱诱导的原生质体膨胀过程中,乙酰胆碱为受体接受后的信号转导可能涉及到Ca2+和CaM,因为Ca2+通道抑制剂硝苯地平(nifedipine, NIF)和La3+可以完全抑制乙酰胆碱诱导的原生质体在含Ca2+介质中的膨胀。同样,钙调素的抑制剂和G蛋白的抑制剂也有这样的作用,而这些化合物在含Na+或K+的介质中则没有作用。
幼叶展开 生长于黑暗中8 d的小麦幼苗,其初生叶的展开受控于光敏色素系统。如果介质中含有Ca2+,乙酰胆碱在暗中可以刺激离体叶切段中幼叶的展开。在没有Ca2+而有Na+的介质中乙酰胆碱也可以刺激黄化小麦初生叶片的展开。在乙酰胆碱的各种衍生物中只有氨基甲酰乙酰胆碱可以刺激黄化小麦初生叶片的展开。乙酰胆碱受体的拮抗剂,阿托品和D-管箭毒可以分别抵消乙酰胆碱在含Ca2+和Na+介质中诱导叶片的展开。乙酰胆碱受体的激活剂,毒蕈碱和烟碱可以分别在Ca2+和Na+的介质中刺激原生质体的膨胀。乙酰胆碱诱导的Ca2+依赖的叶片开展可为Ca2+通道抑制剂硝苯地平和钙调素抑制剂3-氟-甲基吩噻嗪(trifluoperazine, TFP)所减弱,其中只有钙调素抑制剂TFP可以抑制乙酰胆碱诱导的在含Na+介质中黄化小麦初生叶片的展开。
根据以上结果可以初步认为,在植物中乙酰胆碱可能以一种类似于在动物中的机制发挥作用。乙酰胆碱首先与M型和N型乙酰胆碱受体结合。与在动物细胞中一样,M型乙酰胆碱受体可能与磷酸肌醇代谢途径有关,在此途径中,G蛋白,Ca2+通道和钙调素等相继被激活,最后发生生理反应。N型受体是非磷酸肌醇依赖的,它直接控制膜对离子的通透性。这两条途径可以相互独立地引起原生质体的膨胀或叶片的张开。
酶活性乙酰胆碱在植物中的作用机理除参与调节膜对离子的通透性外,可能还涉及对植物体内某些酶活性的调控。乙酰胆碱对兵豆(Lens culinaris)根生长的抑制作用与体内过氧化物同工酶的活性变化密切相关,它可以刺激某些同工酶的活性而抑制另外一些同工酶的活性。
乙酰胆碱本身对于植物体内苯丙氨酸氨基裂解酶的活性和类黄酮的合成没有影响,但它却可以抵消红光对苯丙氨酸氨基裂解酶活性和类黄酮合成的刺激作用。
对内源生长调节物质的影响 乙酰胆碱可以影响植物体内吲哚乙酸和乙烯的代谢。在大豆下胚轴中,乙酰胆碱抑制吲哚乙酸刺激的乙烯合成并抵消它对大豆下胚轴弯钩伸直的抑制作用,它也可以抵消乙烯刺激的蕨类植物原丝体的生长。乙酰胆碱的这种作用可能是通过影响内源吲哚乙酸和乙烯的水平而实现的。以离体大豆叶片的实验证明乙酰胆碱可以抑制组织中乙烯的合成。
乙酰胆碱还可能与内源的赤霉素相互作用。它可以部分代替赤霉素诱导黄瓜下胚轴的伸长,还可以引起植物体内游离态的赤霉素含量增高,这种增高可以阿托品抵消。
生理活动影响中枢胆碱能系统与学习、记忆密切相关,乙酰胆碱(ACh)是中枢胆碱能系统中重要的神经递质之一,其主要功能是维持意识的清醒,在学习记忆中起重要作用。
认知活动关系脑内细胞外乙酰胆碱(ach)的变化主要反映胆碱能神经元的活动,皮层和海马等脑区的ach主要来源于基底前脑胆碱能神经元的纤维投射。应用微透析等技术在体检测清醒、自由活动动物认知过程中脑内乙酰胆碱的含量,可以研究ach与特定行为反应和认知活动之间的关系。研究发现当机体需要对新刺激进行分析时,在学习与记忆、空间工作记忆、注意、自发运动和探究行为等认知活动中,基底前脑胆碱能神经元被激活,脑内ach的释放也随之改变。结果提示脑内胆碱能递质系统活动与认知过程密切相关。
帕金森病关系帕金森病又称"震颤麻痹",是一种中枢神经系统变性疾病,主要是因位于中脑部位"黑质"中的细胞发生病理性改变后,多巴胺的合成减少,抑制乙酰胆碱的功能降低,则乙酰胆碱的兴奋作用相对增强。两者失衡的结果便出现了"震颤麻痹"。
黑质细胞发生变性坏死的原因迄今尚未明了, 可能与遗传和环境因素有关。有学者认为蛋白质、水果、乳制品等摄入不足,嗜酒、外伤、过度劳累及某些精神因素等,均可能是致病的危险因素。 原因不明的多巴胺减少导致的震颤麻痹,在医学上称为"原发性震颤麻痹",即帕金森病;
人体利用人的脑组织有大量乙酰胆碱,但乙酰胆碱的含量会随着年龄的增加而下降。正常老人比青年时下降30%,而老年痴呆患者下降更为严重,可达70%~80%。美国医生伍特曼观察到老年人脑组织乙酰胆碱减少,就给老年人吃富含胆碱的食品,发现有明显的防止记忆减退的作用。英国和加拿大等国的科学家也相继进行了研究,一致认为只要有控制地供给足够的胆碱,可避免60岁左右老年人记忆力减退。所以保持和提高大脑中乙酰胆碱的含量,是解决记忆力下降的根本途径。在自然界是,乙酰胆碱多以胆碱的状态存在于蛋、鱼、肉、大豆等之中,这些胆碱必须在人体内起生化反应后,才能合成具有生理活性的乙酰胆碱。另外,经常服用蜂王浆可以提高脑内乙酰胆碱的含量,从而促进激活脑神经传导功能,提高信息传递速度,增强大脑记忆能力,全面改善脑功能,并能延缓衰老。2
药理作用心血管系统心血管系统ACh对心血管系统主要产生以下作用:
(1)血管扩张作用:静注小剂量本品可由于全身血管扩张而造成血压短暂下降,并伴有反射性心率加快。ACh可引起许多血管扩张。如肺和冠状血管。其扩血管作用主要由于激动血管内皮细胞M,胆碱受体亚型,导致内皮依赖性舒张因子(EDRF)即一氧化氮(nitric oxide,No)释放,从而引起邻近平滑肌细胞松弛,也可能通过压力感受器或化学感受器反射引起。如果血管内皮受损,则ACh的上述作用将不复存在,相反可引起血管收缩。此外,ACh通过激动交感神经末梢突触前膜M1受体,抑制去甲肾上腺素能神经末梢释放NA也与ACh扩血管作用有关。
(2)减慢心率:亦称负性频率作用。ACh能使窦房结舒张期自动除极延缓,复极化电流增加,使动作电位到达阈值的时间延长,导致心率减慢。
(3)减慢房室结和普肯耶纤维传导:即为负性传导作用。ACh可延长房室结和普肯耶纤维(Βurkinje fibers)的不应期,使其传导减慢。当使用强心苷使迷走神经张力增高或全身给药法使用大剂量胆碱受体激动药时所出现的完全性心脏传导阻滞常与房室结传导明显抑制有关。
(4)减弱心肌收缩力:即为负性肌力作用。一般认为胆碱能神经主要分布于窦房结、房室结、普肯耶纤维和心房,而心室较少有胆碱能神经支配,故ACh对心房收缩的抑制作用大于心室。但由于迷走神经末梢与交感神经末梢紧密相邻,迷走神经末梢所释放的ACh可激动交感神经末梢突触前M胆碱受体,反馈性抑制交感神经末梢去甲肾上腺素释放。使心室收缩力减弱。
(5)缩短心房不应期:ACh不影响心房肌的传导速度,但可使心房不应期及动作电位时程缩短(即为迷走神经作用)。
胃肠道ACh可明显兴奋胃肠道平滑肌,使其收缩幅度、张力、蠕动增加,并可促进胃、肠分泌,引起恶心、嗳气、呕吐、腹痛及排便等症状。
泌尿道ACh可使泌尿道平滑肌蠕动增加,膀胱逼尿肌收缩,使膀胱最大自主排空压力增加,降低膀胱容积,同时膀胱三角区和外括约肌舒张,导致膀胱排空。
其他(1)腺体:ACh可使泪腺、气管和支气管腺体、唾液腺、消化道腺体和汗腺分泌增加。
(2)眼:当ACh局部滴眼时,可致瞳孔收缩,调节于近视。
(3)神经节和骨骼肌:ACh作用于自主神经节NN胆碱受体和骨骼肌神经肌肉接头的NM胆碱受体,引起交感、副交感神经节兴奋及骨骼肌收缩。此外,因肾上腺髓质受交感神经节后纤维支配,故NN胆碱受体激动能引起肾上腺素释放。
(4)中枢:由于ACh不易进人中枢,故尽管中枢神经系统有胆碱受体存在,但外周给药很少产生中枢作用。
(5)支气管:ACh可使支气管收缩。
(6)ACh还能兴奋颈动脉体和主动脉体化学受体。3
本词条内容贡献者为:
王强 - 副教授 - 西南大学
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国