

海洋细菌是生活在海洋中的、不含叶绿素和藻蓝素的原核单细胞生物。它们是海洋微生物中分布最广、数量最大的一类生物,个体直径常在1微米以下,呈球状、杆状、螺旋状和分枝丝状的微生物
概述生活在海洋中的、不含叶绿素和藻蓝素的原核单细胞生物。它们是海洋微生物中分布最广、数量最大的一类生物,个体直径常在1微米以下,呈球状、杆状、螺旋状和分枝丝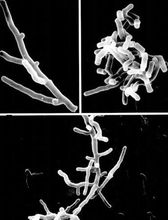 状的微生物。无真核、细胞壁坚韧。能游动的种以鞭毛运动。严格地说,海洋细菌是指那些只能在海洋中生长与繁殖的细菌。
状的微生物。无真核、细胞壁坚韧。能游动的种以鞭毛运动。严格地说,海洋细菌是指那些只能在海洋中生长与繁殖的细菌。
19世纪中期首次分离出一个海洋细菌,1865年分离出其中的奇异贝氏硫细菌。从1884年起,又研究深海细菌。早期只注重分类,1946年后进入以研究其生理和生态为基础的阶段。
海洋细菌有自养和异养、光能和化能、好氧和厌氧、寄生和腐生以及浮游和附着等不同类型。海水中以革兰氏阴性杆菌占优势,常见的有假单胞菌属、弧菌属、无色杆菌属、黄杆菌属、螺菌属、微球菌属、八叠球菌属、芽孢杆菌属 、棒杆菌属、枝动菌属、诺卡氏菌属和链霉菌属等10多个属;洋底沉积物中以革兰氏阳性细菌居多;大陆架沉积物中以芽孢杆菌属最常见。1
研究简史19世纪中期,有人就分离出第一个海洋细菌,1865年又分离出海洋奇异贝氏硫细菌。深海细菌的研究也于1884年开始。但在相当长的时间内,一直停留在描述、分类的水平上。1946年,美国C.E.佐贝尔以海洋细菌为主要内容的《海洋微生物学》一书的问世,促使海洋微生物的研究进入以生理、生态为基础的阶段。
1959年以后,苏联学者A.E.克里斯连续出版了研究深海微生物的著作,提出微生物海洋学的研究设想。1961年国际海洋微生物学讨论会的召开,标志着以海洋细菌为主要内容的海洋微生物学已成为独立的学科。60年代以来,代表性的专著有美国学者E.J.F.伍德1965年出版的《海洋微生物生态学》,J.M.西伯斯1979年出版的《海洋微生物》等。
种类组成海洋中有自养和 异养、光能和化能、好氧和厌氧、寄生和腐生以及浮游和附着等类型的细菌。几乎所有已知生理类群的细菌,都可在海洋环境中找到。最常见的有:
异养、光能和化能、好氧和厌氧、寄生和腐生以及浮游和附着等类型的细菌。几乎所有已知生理类群的细菌,都可在海洋环境中找到。最常见的有:
假单胞菌属 (Pseudomonas)、弧菌属(Vibrio)、无色杆菌属 (Achromobacter)、黄杆菌属(Flavobacterium)、螺菌属(Spirillum)、微球菌属(Micrococcus)、八叠球菌属(Sarcina)、芽孢杆菌属(Bacillus)、棒杆菌属 (Corynebacterium)、枝动菌属(Mycoplana)、诺卡氏菌属(Nocardia)和链霉菌属(Streptomyces)等十多个属。在海水中,革兰氏阴性杆菌占优势;在远洋沉积物中,则革兰氏阳性细菌居多;在大陆架沉积物中,芽孢杆菌属最为常见。2
生态分布海洋细菌在海洋中分布广、数量多,是海洋微生物中最重要的成员。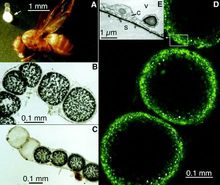 其数量分布特点是,近海区的细菌密度较远洋区大,尤以内湾和河口区最大。每毫升近岸海水中一般可分离到102~103个细菌菌落,有时超过105个;而在每毫升深海海水中,有时却分离不出一个细菌菌落。
其数量分布特点是,近海区的细菌密度较远洋区大,尤以内湾和河口区最大。每毫升近岸海水中一般可分离到102~103个细菌菌落,有时超过105个;而在每毫升深海海水中,有时却分离不出一个细菌菌落。
表层海水和水-底泥界面处的细菌密度较深层水大,底泥中的细菌密度一般较海水中大,泥土底质中的细菌密度一般高于沙土底质。在每克底泥中细菌数量约在102~105个之间,高的可达到106个以上。在海洋调查中,有时发现某水层中的细菌数量剧增,出现不均匀的微分布现象。这种现象主要是由于海水中可供细菌利用的有机物质分布不均匀所引起,一般在赤潮之后常伴随着细菌数量的剧增。
生活习性陆源细菌可在近岸海域和远洋上水层区中适应生存,因此生活在这些环境中 的海洋细菌的生理生态特性与陆生细菌相似。但生活在深海的细菌,因深海环境具有高盐、高压、低温和低营养等特点,因此其生理、生态特性与陆源细菌迥然不同。
的海洋细菌的生理生态特性与陆生细菌相似。但生活在深海的细菌,因深海环境具有高盐、高压、低温和低营养等特点,因此其生理、生态特性与陆源细菌迥然不同。
嗜盐性海洋细菌最普遍的特性。海水中含有各种盐类和微量元素,如钠是海洋细菌生长所必需的,在输送基物进入细胞的过程中起着不可取代的作用。此外,钾、镁、钙、磷、硫和其他微量元素也是某些海洋细菌生长所必需的。
嗜冷性绝大多数海洋细菌都具有在低温下生长的特性。在海洋中,90%以上水体的温度是在5℃以下。海洋中的高温细菌,只在海底热泉的特异环境中发现过。某些中温细菌,虽然其最适生长温度为20℃左右,但它也能在0℃下缓慢生长,这些细菌称为耐低温细菌。那些在0℃或0℃以下生长良好的细菌,称为嗜冷细菌。嗜冷细菌对热反应极为敏感,20~25℃的中温已足以阻碍其生长与代谢,它们的细胞膜构造具有适应低温的特点,主要分布于极地、深海和高纬度的海洋中。
嗜压性海洋中水深每增加10米,静水压力便递增1大气压。海洋最深处,静水压力可超过1000大气压;整个海洋约有一半水深超过3800米,静水压力为380~1100大气压,这种压力妨碍了浅海和陆源细菌在深海中的生长。深海嗜压细菌具有适应高压而生长代谢的能力,并能在高压环境中保持酶系统的稳定性。
低营养性深海中营养物质较为稀少,某些浮游型的海洋细菌适应于低浓度营养的海水。因此,分离培养海洋细菌忌用营养丰富的培养基。20世纪60年代以来,美国学者H.W.詹纳施采用连续培养技术,在恒化器中研究海洋细菌对低营养的反应、对有机化合物的吸收与利用等,取得较接近于海洋中实际状况的成果。
趋化性与附着生长海水中营养物质虽然较为稀少,但海洋中各种固体表面和不同性质的界 面上,却吸附积聚着较为丰富的营养物。绝大多数海洋细菌都具有运动能力,某些细菌还具有沿着某种化合物的浓度梯度而移动的能力,这一特点称为趋化性。某些专门附着于海洋植物体表面而生活的细菌,称为附生植表细菌。海洋细菌附着生长的特性,对于海洋生物和非生物固体表面膜的形成起着重要作用,如海洋物体表面污着生物的形成,就是在这个基础上发展起来的(海洋污着生物)。
面上,却吸附积聚着较为丰富的营养物。绝大多数海洋细菌都具有运动能力,某些细菌还具有沿着某种化合物的浓度梯度而移动的能力,这一特点称为趋化性。某些专门附着于海洋植物体表面而生活的细菌,称为附生植表细菌。海洋细菌附着生长的特性,对于海洋生物和非生物固体表面膜的形成起着重要作用,如海洋物体表面污着生物的形成,就是在这个基础上发展起来的(海洋污着生物)。
多形性有的菌株在培育中可以观察到多种形态,包括球形、椭球形、大小不一的杆状或各种不规则形态的细胞。
发光性少数属类具有发光特性。
另外,海洋细菌的多态性特点也是常见的。在同一株细菌的纯培养中,往往可观察到多种形态的细胞。少数几个海洋细菌属还具有发光的特性,发光细菌通常可以从海水和新鲜的鱼虾体上分离到。2
生态作用在海洋生态系中的作用海洋经历着剧烈的变动而又不断地保持着动态平衡,始终富有生命力和生产力,海洋细菌在其中起着重要的作用。当海洋生态系的动态平衡遭受某种破坏时,海洋细菌以其敏感的适应能力和极快的繁殖速度,迅速形成异常微生物区系,积极参与氧化、还原活动,调整和促进新动态平衡的形成和发展。
在海洋氮循环中的作用海洋氮循环的基本途径与陆地相仿,至今尚未从海洋中直接分离得到根瘤菌,但通过定量PCR方法发现地中海腐殖泥中有大量放射型根瘤菌(Rhizobium radiobacter)。固氮菌可以从海洋中分离到,硝化细菌多集中分布于海洋沉积物中。在海水中,硝酸盐的含量随着靠近海底沉积物的距离而逐渐增加,因此硝化作用在大陆架和近岸海域较为明显,海洋中的硝酸盐主要是通过这一途径产生。反硝化作用在有机物来源丰富、溶解氧浓度低的内湾和河口海域较为强烈,反硝化细菌在一定条件下影响海洋中可利用状态的氮。
在海洋硫循环中的作用某些异养细菌分解含硫蛋白类物质时产生硫化氢;在有机物丰富的浅海嫌气水域,硫酸盐还原细菌还原硫酸盐时,也产生大量硫化氢,污染大片海湾与滩涂。这些硫化氢可由各种硫细菌逐步氧化,最终形成硫酸盐。
在海洋磷循环中的作用细菌分解海洋动植物残体,并释放出可供植物利用的无机态磷酸盐。磷也是海洋微生物繁殖和分解有机物过程所必需的因子。
在海洋食物链中的作用海洋细菌多数是分解者,有一部分是生产者,因而具有双重性,参与海洋物质分 解和转化的全过程。在嫌气条件下,有机物质分解的最终产物是甲烷和硫化氢等;在多氧条件下,有机物质的分解是不完全的。在海洋中,分解有机物的代表性菌群是随着被作用有机物的类别而不同的:分解有机含氮化合物者,分别有液化明胶、消化鱼蛋白、蛋白胨多肽、氨基酸、含硫蛋白以及分解尿素等细菌;分解碳水化合物者,分别有分解各种糖类、淀粉、纤维素、琼胶、褐藻酸以及甲壳素等细菌。另有降解烃类化合物以及利用芳香化合物(如酚等)的细菌。海洋细菌分解有机物质的终极产物,如氨、硝酸盐、磷酸盐以及二氧化碳等,都直接或间接地为海洋植物提供营养。可见,它们在海洋无机营养再生过程中起着重要的作用。海洋细菌自身增殖的生物量,也为海洋原生动物、浮游动物以及底栖动物等提供直接的营养。除异养细菌外,某些海洋细菌具有光合作用的能力。另一类海洋化能自养的细菌,从氧化氨、硝酸盐、甲烷、分子氢和硫化氢等中,取得能量而增殖。如在海底热泉的特殊生态系中,硫磺细菌是利用硫化氢作为能源而进行有机碳的初级生产。由于深海是非光合作用区,其生物区系的构成有其独特性,因此细菌在深海生态系食物链中的作用,已引起重视。
解和转化的全过程。在嫌气条件下,有机物质分解的最终产物是甲烷和硫化氢等;在多氧条件下,有机物质的分解是不完全的。在海洋中,分解有机物的代表性菌群是随着被作用有机物的类别而不同的:分解有机含氮化合物者,分别有液化明胶、消化鱼蛋白、蛋白胨多肽、氨基酸、含硫蛋白以及分解尿素等细菌;分解碳水化合物者,分别有分解各种糖类、淀粉、纤维素、琼胶、褐藻酸以及甲壳素等细菌。另有降解烃类化合物以及利用芳香化合物(如酚等)的细菌。海洋细菌分解有机物质的终极产物,如氨、硝酸盐、磷酸盐以及二氧化碳等,都直接或间接地为海洋植物提供营养。可见,它们在海洋无机营养再生过程中起着重要的作用。海洋细菌自身增殖的生物量,也为海洋原生动物、浮游动物以及底栖动物等提供直接的营养。除异养细菌外,某些海洋细菌具有光合作用的能力。另一类海洋化能自养的细菌,从氧化氨、硝酸盐、甲烷、分子氢和硫化氢等中,取得能量而增殖。如在海底热泉的特殊生态系中,硫磺细菌是利用硫化氢作为能源而进行有机碳的初级生产。由于深海是非光合作用区,其生物区系的构成有其独特性,因此细菌在深海生态系食物链中的作用,已引起重视。
在海洋动物植物体表和动物消化道内,存在着特异的微生物区系,如孤菌等是海洋动物消化道中常见的细菌;分解甲壳素的细菌也能在肉食性动物消化道中发现;利用各种多糖类的细菌常是海藻体表的重要菌群,如褐藻酸降解菌是海带上的优势菌群。细菌的中间代谢产物,如抗菌素、氨基酸、维生素和毒素等,是促进或限制某种海洋生物生存和生长的因素。某些浮游生物与细菌之间存在着相互依存的营养关系,如细菌为浮游植物提供必需的维生素等营养物质,浮游植物分泌乙醇酸等物质为某些细菌提供能源和碳源。
研究意义海洋细菌参与降解各种海洋污染物和毒物的过程,有助于保持海洋生态系的平衡和促进海洋自净能力;海洋细菌是产生新抗菌素、氨基酸、维生素和其他生理活性物质的重要生产者;细菌参与海洋的沉积成岩作用,如参与硫矿和深海锰结核的形成等;在海洋成油、成气的过程中,细菌起着重要作用;海水具有杀菌效果,是由于海洋细菌的拮抗和溶菌作用,致使陆源致病菌迅速死亡;海洋细菌可直接作为海洋经济动物的饵料;细菌参与对各种海洋物质的腐蚀、变性、污秽和破坏过程;某些海洋细菌是人体或海洋生物的致病菌;在特定条件下,海洋细菌代谢产物的积累会毒化养殖环境,如氨和硫化氢的积累危害生物养殖;也可以利用细菌的代谢活动来改善被毒化的养殖环境,如氨的氧化等。3
国内研究状况解放前,中国海洋细菌研究几乎是空白。解放后,随着海洋科学的发展,海洋细菌研究日益受到重视。1956年中国科学院海洋生物研究室(中国科学院海洋研究所前身)正式成立了海洋细菌组。两年后,山东海洋学院成立,海洋生物系与水产系也开展了海洋细菌方面的工作。到60年代中期,主要进行了海洋调查及分类方面研究。1959年,薛廷跃、孙国玉等首次从青岛近岸潮间带底泥中分离出排流杆菌、氧化硫杆菌及脱氧硫杆菌。1960年,薛廷跃从胶州湾海水中获得11株小珠菌,它们是丹砂色小珠菌共5株,StrainA一E,硫色小球菌2株,StrainA一B,柠檬色小球菌海洋变种、李氏小球菌海洋变种和纯白色小球菌。1961年,陈世阳等从青岛近海分离获得嗜盐性自身固氮菌海洋变种。之后研究逐渐扩大,研究厌氧细菌分离,从胶州湾海泥中获得厌氧光合细菌——纤细红硫菌。70年代中期后10年左右的时间,随着黄岛输油码头的建立,青岛近岸油污事故时有发生,同时胶州湾养殖藻类病害也频频出现。因此,这期间着重开展了环保及与藻类病害有关的研究工作。1979年丁美丽等在国内首次报道有关石油烃降解菌生态研究结果,从胶州湾分离出300余株具有分解石油烃能力的微生物。其中,有5株放线菌,1株酵母菌,其余都是细菌,它们隶属于假单孢杆菌属、不动杆菌属、短杆菌属、产碱杆菌属、棒状菌属、节细菌属、弧菌属,另外还发现有肠道杆菌科及类似奈瑟氏球菌属的革兰氏阴性球菌。1983年,沈世泽等在青岛近海发现有还原络细菌存在。1986年,陈騳经调查胶州湾或海带栽培区在异养菌中褐藻酸降解菌有相当高的比例。1979年和1990年,陈騳等和丁美丽先后在海带病害研究中发现海带病烂与褐藻酸降解菌有关。
进入80年代后,中国对虾养殖业迅速发展。对虾疾病频繁发生,有关单位先后开展养殖动物病害研究,并继续进行有关环境保护工作。1983年,王文兴等从青岛太平角及即墨沿海养殖场水样及泥样中分离出弧菌属、假单孢杆菌属、不动杆菌属、棒状杆菌属、微球菌等属的细菌,还从对虾体内分离出一批菌株。它们分别属于弧菌属、棒状杆菌属、假单孢杆菌属、不动杆菌属、产碱杆菌和副溶血性弧菌。
此外,钱振懦于1985年发现在青岛近岸水生活着许多对污水细菌具有抑制作用的细菌,如苛求芽孢杆菌、短芽孢杆菌、球形芽孢杆菌、坚强芽孢杆菌、朱红微球菌、李氏微球菌、亮白微球菌和放线菌。徐怀恕等于1989年发现在青岛港钢板桩上附着对钢有腐蚀作用的氧化硫杆菌、排硫杆菌和脱氮杆菌。4
本词条内容贡献者为:
郭亮 - 副教授 - 中国海洋大学
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国