

MicroRNAs(miRNAs)是在真核生物中发现的一类内源性的具有调控功能的非编码RNA,其大小长约20~25个核苷酸。成熟的miRNAs是由较长的初级转录物经过一系列核酸酶的剪切加工而产生的,随后组装进RNA诱导的沉默复合体(RNA-induced silencing complex,RISC),通过碱基互补配对的方式识别靶mRNA,并根据互补程度的不同指导沉默复合体降解靶mRNA或者阻遏靶mRNA的翻译。最近的研究表明miRNA参与各种各样的调节途径,包括发育、病毒防御、造血过程、器官形成、细胞增殖和凋亡、脂肪代谢等等。
简介MicroRNA (miRNA):
 是含有茎环结构的miRNA前体,经过Dicer加工之后的一类非编码的小RNA分子(18~25个核苷酸)。MiRNA,以及miRISCs(RNA诱导基因沉默复合物)在动物和植物中广泛表达。因之具有抑制靶mRNA转录、翻译或者能够剪切靶mRNA并促进其降解的功能,miRNA被认为在调控发育过程中有重要作用。1
是含有茎环结构的miRNA前体,经过Dicer加工之后的一类非编码的小RNA分子(18~25个核苷酸)。MiRNA,以及miRISCs(RNA诱导基因沉默复合物)在动物和植物中广泛表达。因之具有抑制靶mRNA转录、翻译或者能够剪切靶mRNA并促进其降解的功能,miRNA被认为在调控发育过程中有重要作用。1
microRNAs(miRNAs)是一种小的,类似于siRNA的分子,由高等真核生物基因组编码,miRNA通过和靶基因mRNA碱基配对引导沉默复合体(RISC)降解mRNA或阻碍其翻译。miRNAs在物种进化中相当保守,在植物、动物和真菌中发现的miRNAs只在特定的组织和发育阶段表达,miRNA的组织特异性和时序性,决定组织和细胞的功能特异性,表明miRNA在细胞生长和发育过程中起多种调节作用。
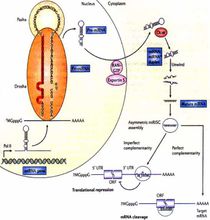
作用机理miRNA基因通常是在核内由RNA聚合酶II(pol II)转录的,最初产物为大的具有帽子结构(7MGpppG)和多聚腺苷酸尾巴(AAAAA)的pri-miRNA。pri-miRNA在核酸酶Drosha和其辅助因子Pasha的作用下被处理成70个核苷酸组成的pre-miRNA。RAN–GTP和exportin 5将pre-miRNA输送到细胞质中。随后,另一个核酸酶Dicer将其剪切产生约为22个核苷酸长度的miRNA:miRNA*双链。这种双链很快被引导进入沉默复合体(RISC)复合体中,其中一条成熟的单链miRNA保留在这一复合体中。成熟的miRNA结合到与其互补的mRNA的位点通过碱基配对调控基因表达。
与靶mRNA不完全互补的miRNA在蛋白质翻译水平上抑制其表达(哺乳动物中比较普遍)。然而有证据表明,这些miRNA也有可能影响mRNA的稳定性。使用这种机制的miRNA结合位点通常在mRNA的3’端非编码区。如果miRNA与靶位点完全互补(或者几乎完全互补),那么这些miRNA的结合往往引起靶mRNA的降解(在植物中比较常见)。通过这种机制作用的miRNAs的结合位点通常都在mRNA的编码区或开放阅读框中。每个miRNA可以有多个靶基因,而几个miRNAs也可以调节同一个基因。这种复杂的调节网络既可以通过一个miRNA来调控多个基因的表达,也可以通过几个miRNAs的组合来精细调控某个基因的表达。随着miRNA调控基因表达的研究的逐步深入,将帮助我们理解高等真核生物的基因组的复杂性和复杂的基因表达调控网络。
特点广泛存在于真核生物中, 是一组不编码蛋白质的短序列RNA , 它本身不具有开放阅读框架(ORF) ;
70 %的哺乳动物miRNA 基因是位于TUs区
通常的长度为18~25 nt , 但在3′端可以有1~2 个碱基的长度变化;
成熟的miRNA 5′端有一磷酸基团, 3′端为羟基, 这一特点使它与大多数寡核苷酸和功能RNA 的降解片段区别开来;
多数miRNA 还具有高度保守性、时序性和组织特异性。
生理作用在正常生理过程中的作用
只有一小部分miRNAs生物学功能得到阐明。这些miRNAs调节了细胞生长,组织分化,因而与生命过程中发育、疾病有关。通过对基因组上miRNA的位点分析,显示其在发育和疾病中起了非常重要的作用。一系列的研究表明:miRNAs在细胞生长和凋亡,血细胞分化,同源异形盒基因调节,神经元的极性,胰岛素分泌,大脑形态形成,心脏发生,胚胎后期发育等过程中发挥重要作用。例如,miR-273和lys-6编码的miRNA,参与线虫的神经系统发育过程;miR-430参与斑马鱼的大脑发育;miR-181控制哺乳动物血细胞分化为B细胞;miR-375调节哺乳动物胰岛细胞发育和胰岛素分泌;miR-143在脂肪细胞分化起作用;miR-196参与了哺乳动物四肢形成,miR-1与心脏发育有关。另有研究人员发现许多神经系统的miRNAs在大脑皮层培养中受到时序调节,表明其可能控制着区域化的mRNA翻译。对于新的miRNA基因的分析,可能发现新的参与器官形成、胚胎发育和生长的调节因子,促进对癌症等人类疾病发病机制的理解。
与癌症最近的研究发现,miRNA表达与多种癌症相关,大约50%得到注解的miRNAs在基因组上定位于与肿瘤相关的脆性位点(fragile site)。这说明miRNAs在肿瘤发生过程中起至关重要的作用,这些miRNAs所起的作用类似于抑癌基因和癌基因的功能,有研究人员将miRNA命名为“oncomirs”。
充当抑癌基因作用的miRNA:
mir-125b-1,位于染色体的11q24脆性位点,乳腺癌、肺癌、卵巢癌、子宫癌病人中11q924位点常有缺失,而这一位点并不存在已知的抑癌基因。
65%B细胞慢性淋巴型白血病(CLL)病人,50%的套细胞淋巴瘤病人,16-40%的骨髓瘤病人、60%的前列腺癌病人中有13q14位点的缺失。因此,在这个30Kb的区域中必定有一个肿瘤抑制基因的存在。而mir-15a和mir-16-1位于这一区域中一个功能未知的被称为LEU2的非编码蛋白的RNA基因的内含子区域。Climmino等报道,miR-15a和miR-16-1负调控BCL2,一个抗凋亡基因,因此,这两个miRNAs的缺失或下调,导致了BCL2表达的升高,促进了白血病、淋巴瘤和前列腺癌的发生。
有研究报道,mir-143和mir-145在结肠癌中明显下调。有趣的是,其发夹结构的前体分子在肿瘤和正常组织中含量相似,这表明,可能是由于其成熟过程受到破坏。mir-143和mir-145的肿瘤抑制基因功能不仅仅局限于结肠癌,在乳腺癌、前列腺癌、子宫癌、淋巴癌等细胞系中其表达量也明显下调。
Takamizawa等发现肺癌病人的let-7表达显著降低,并且这导致这些病人更差的预后,非小细胞肺癌病人的let-7表达水平越低,其预后越差,术后生存期越短。体外组织培养实验表明,在人的肺癌细胞中瞬时的表达let-7可以抑制细胞的增殖,这也说明let-7在肺组织中可能是一个抑癌基因。实验表明,在人类细胞中let-7通过3’非翻译区域直接抑制癌基因Ras的表达。大约有15-30%的人类肿瘤都含有Ras突变,而激活的突变导致这个蛋白表达上升可以引起细胞转化。因此,能够调节Ras蛋白表达的let-7,可以控制细胞的增殖速度。
充当癌基因作用的miRNA:
miR-21在胶质母细胞瘤中表达增加。这个基因在肿瘤组织中表达量比正常组织高5-100倍。反义核酸的研究发现这个miRNA通过抑制凋亡而并非影响细胞增殖控制细胞生长,这预示着这个miRNA具有癌基因的功能。另一项独立研究,利用芯片来检测肿瘤和正常组织的245个miRNAs的表达水平,也发现胶质母细胞瘤中miR-21表达升高。由于mir-21不是一个大脑特异性的基因,在乳腺癌样品中表达也有增加,这个基因可能在肿瘤发生中起到广泛的作用。
Metzler等人发现,在BIC基因上具有一段138个核苷酸的保守序列,编码mir-155的发夹结构。该研究组还发现在Burkitt淋巴瘤中,miR-155表达量上升了100倍,此外的研究也发现,Hodgkin淋巴瘤等肿瘤中miR-155水平也有提高。因此,mir-155可能是作为一个癌基因和MYC协同作用,而其正常功能是在B细胞的分化中起作用,其可能的靶基因是那些对抗MYC信号通路的基因。
He等人发表的论文发现在散布的B细胞淋巴瘤、滤泡型淋巴瘤、套细胞淋瘤等肿瘤中常常有13q31位点的扩增。而在这一扩增区域的唯一的基因就是一个非编码蛋白的RNA,C13orf25,这个转录本编码了mir-17-92基因簇,其中包含了7个miRNAs:miR-17-5p,miR-17-3p,miR-18a,miR-19a,miR-20a,miR-19b-1,and miR-92-1。他们由此推断,这个基因簇的过表达与肿瘤形成有关,而后通过实验研究证明,过表达Myc和mir-17-19-b1基因簇淋巴癌与仅有Myc过表达肿瘤相比较,具有更强的增殖能力,更低的细胞死亡率。这些实验证实,mir-17-19-b1中的miRNAs可以协同地行使癌基因的功能,其靶基因可能是在MYC过表达条件下激活的凋亡蛋白。当凋亡途径被mir-17-19-b1去除,MYC可以诱导细胞不受控制地增殖,这导致了肿瘤发生。
O’Donnell等人单独证明了mir-17-92基因簇是一组可能的肿瘤相关的基因。他们用miRNAs芯片筛选过表达MYC,B细胞系P493-6中miRNA表达变化。他们发现MYC诱导了mir-17-92的表达,而这些miRNAs可以抑制E2F1的翻译。在这一模型中,mir-17-92基因簇所起的作用似乎是抑癌基因的作用,这与上面讨论的He等人的发现是相反的。这其中可能的机理是,尽管E2F1可以促进细胞增殖,但是当E2F1的表达水平超过一阈值,它也可以引起凋亡,在此情况下,miRNAs对于E2F1的负调控可能是通过阻断E2F1的诱导凋亡活性,从而促进MYC介导的细胞增殖,支持了He等提出的模型。
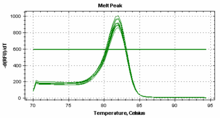
检测手段迄今为此,对miRNA 的检测方法主要有Northern Blot 等基于分子杂交的方法,这些方法敏感度低、耗时长、RNA 的用量较大。国际上对于miRNA进行检测,以PCR 技术来对miRNA 进行检测作为验证标准,其具有快速、特异性强、灵敏度高等优点。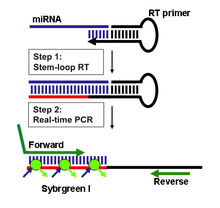 分为欧米伽引物检测法和实时荧光定量PCR两大类,欧米伽引物检测法可以达到对miRNA拷贝数的绝对定量,RT反应采用欧米伽引物,样品miRNA和内置的4个标准在同一反应管中进行同步RT、PCR,最后进行基因片段分析,通过对比实测的标准曲线而做到绝对定量。
分为欧米伽引物检测法和实时荧光定量PCR两大类,欧米伽引物检测法可以达到对miRNA拷贝数的绝对定量,RT反应采用欧米伽引物,样品miRNA和内置的4个标准在同一反应管中进行同步RT、PCR,最后进行基因片段分析,通过对比实测的标准曲线而做到绝对定量。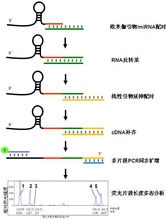 实时荧光定量PCR分两种:Taqman探针法和SYBR Green荧光染料法。 检测原理:茎环结构的逆转录引物能与miRNA 3'端部分结合,在逆转录酶的作用下进行逆转录反应,然后特异的正反向引物和SYBRGreen荧光染料(或探针)共同参与的定量PCR反应体系,实现对逆转录产物进行定量检测。(1)基于茎环结构引物的逆转录反应;(2) 实时荧光定量PCR。
实时荧光定量PCR分两种:Taqman探针法和SYBR Green荧光染料法。 检测原理:茎环结构的逆转录引物能与miRNA 3'端部分结合,在逆转录酶的作用下进行逆转录反应,然后特异的正反向引物和SYBRGreen荧光染料(或探针)共同参与的定量PCR反应体系,实现对逆转录产物进行定量检测。(1)基于茎环结构引物的逆转录反应;(2) 实时荧光定量PCR。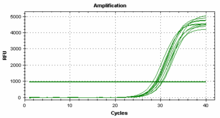
研究手段miRNA鉴定及功能研究手段
鉴定miRNA常用的方法包括直接克隆鉴定,miRNA芯片分析和生物信息学预测。当然计算机预测的miRNA必须经过RT-PCR或Northern试验分析才能鉴定,另外也可以通过miRNA mimics和inhibitors从功能上进行实验鉴定。另外序列分析表明,至少有1/3的人类基因与miRNA调控相关,而且越来越多的试验证据表明miRNA还有很多功能未被发现。研究基因的功能通常是将其从基因组中敲除,然后观察敲除前后的变化,但在破译miRNA的功能,我们一般不会采用这种策略,而是通过增强或减弱该miRNA的表达来鉴定其功能。
miRNA mimics是模拟生物体内源的miRNAs,运用化学合成的方法合成,能增强内源性miRNA的功能。而miRNA inhibitor是化学修饰的专门针对细胞中特异的靶miRNA的抑制剂。
人工合成的miRNA(artificial miRNA,amiRNA)已经成功应用于沉默预期靶基因的表达及其功能研究,人工合成的miRNAs既能够特异性地沉默单一基因,也可以同时沉默多个相关但不相同的基因。miRNA mimics进一步增强内源miRNA的沉默作用,降低细胞内蛋白表达量,进行功能获得性(gain-of-function)研究;相反,使用化学合成的方法合成miRNA inhibitors,特异的靶向和敲除单个的miRNA分子,可以削弱内源miRNA的基因沉默效应,提高蛋白表达量,进行功能缺失性(loss-of -function)研究,可以用来筛选miRNA靶位点,筛选调控某一基因表达的miRNA,筛选影响细胞发育过程的miRNA。化学合成miRNA mimics和inhibitors是研究的一个新热点,已经成为研究动植物基因家族功能的有用工具,并有望成为癌症治疗和临床研究的一种新策略。
miRNA 的特点:MiRNAs具有高度的保守性、时序性和组织特异性。miRNAs的表达方式各不相同。线虫和果蝇当中的部分miRNA在各个发育阶段都有表达而且不分组织和细胞特性,而其他的miRNA则表现出更加严谨的时空表达模式(a more restricted spatial and temporal expression pattern)——只有在特定的时间、组织才会表达。细胞特异性或组织特异性是miRNA的表达的主要特点,又如拟南芥中的miR-171仅在其花序中高水平表达,在某些组织低水平表达,在茎、叶等组织中却无任何表达的迹象;20-24h的果蝇胚胎提取物中可发现miR-12,却找不到miR3-miR6,在成年果蝇中表达的miR-1和let-7也无法在果蝇胚胎中表达,这同时体现了miRNA的又一特点——基因表达时序性。MiRNA表达的时序性和组织特异性提示人们miRNA的分布可能决定组织和细胞的功能特异性,也可能参与了复杂的基因调控,对组织的发育起重要作用。
展望miRNA在细胞分化,生物发育及疾病发生发展过程中发挥巨大作用,越来越多的引起研究人员的关注。随着对于miRNA作用机理的进一步的深入研究,以及利用最新的例如miRNA芯片等高通量的技术手段对于miRNA和疾病之间的关系进行研究,将会使人们对于高等真核生物基因表达调控的网络理解提高到一个新的水平。这也将使miRNA可能成为疾病诊断的新的生物学标记,还可能使得这一分子成为药靶,或是模拟这一分子进行新药研发,这将可能会给人类疾病的治疗提供一种新的手段。
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国