

半索动物又称隐索动物,是一些口腔背面有一条短盲管(口盲囊buccal diverticulum,俗称口索stomochord)前伸至吻内的海栖类群。1825年黄殖翼柱头虫(Pty-choderaflava)最早被发现和命名,全世界至少已发现90余种,包括体呈蠕虫状的肠鳃纲和形似苔藓动物的羽鳃纲两大类,其中有77%以上的种类隶属于肠鳃纲,最常见的代表动物为各种柱头虫。1
本门动物的体长由2.3mm~2.5m不等,大多数种类广泛分布于热带海和温带海的沿海,只有极少数种能生存在寒带海中。主要栖息于潮间带或潮下带的浅海沙滩、泥地或岩石间,营单体自由生活或集群固着生活,40m以下的海域中种类甚少,在西非大西洋4500m深海所发现的粗吻柱头虫(Glandiceps sp.),是迄今所知生活在海底最深的半索动物。1
下属分类分三个纲:肠鳃纲,羽鳃纲,笔石纲。
肠鳃纲柱头虫,黄岛长舌虫(青岛产),为自由运动的种类。是本门的代表动物。1
柱头虫(Balanoglossus)属于肠鳃纲(Enteropneusta),是半索动物门中分布甚广的类群,产于我国的三崎柱头虫(B.misakiansis)具有本门动物的主要特征。体呈蠕虫形,两侧对称而背腹明显,全身由吻(proboscis)、领(collar)和躯干(trunk)三部分组成,吻位于最前端,稍后是指环状的领,躯干部最长,又可分为鳃裂区、生殖区、肝囊区和肠区,末端为肛门。虫体内的各部均有空腔,即由体腔分化而成的吻腔、领腔和躯干腔。柱头虫凭藉富含肌肉和腔内充满海水的圆锥形吻部,在浅海沙滩中运动和挖掘成U字形洞道,并藏身在洞道内营少动的生活,人们可在退潮时于其洞口看到盘曲成条的粪便(图13-2)。1
解剖结构
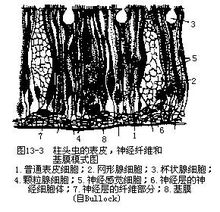 a.体壁和体腔:体壁由表皮、肌肉层和体腔膜构成。表皮的外层是单层较厚的上皮,外被纤毛,除肝囊区外,上皮内含有形状各异的多种腺细胞,均可分泌粘液至体表,粘牢洞道壁上的沙粒,使之不致坍塌。外层下为神经细胞体及神经纤维交织而成的神经层,底部则为薄而无结构的基膜(图13-3)。基膜的深处是环肌、纵肌和结缔组织合成的平滑肌层,紧贴其内的为体腔膜。1
a.体壁和体腔:体壁由表皮、肌肉层和体腔膜构成。表皮的外层是单层较厚的上皮,外被纤毛,除肝囊区外,上皮内含有形状各异的多种腺细胞,均可分泌粘液至体表,粘牢洞道壁上的沙粒,使之不致坍塌。外层下为神经细胞体及神经纤维交织而成的神经层,底部则为薄而无结构的基膜(图13-3)。基膜的深处是环肌、纵肌和结缔组织合成的平滑肌层,紧贴其内的为体腔膜。1
吻内有一吻腔,后背部以吻孔与外界相通,可容水流进入和废液排出,当吻腔充水时,吻部变得坚挺有力,形似柱头,可用于穿洞凿穴,柱头虫即因此而得名。领和躯干部被背、腹隔膜分为成对的领腔及躯干腔,这5个腔都是由体腔分化而来。1
b.消化和呼吸:柱头虫的消化道是从前往后纵贯于领和躯干末端之间的一条直管。口位于吻、领的腹面交界处,口腔背壁向前突出一个短盲管至吻腔基部,盲管的腹侧有胶质吻骨(pro-boscis skeleton),但尚无坚硬结构,因此过去曾被视作雏形脊索而称为口索(stomochord),也有人认为短育管可能是脊椎动物脑垂体前叶的前身。由于口索形甚短小,所以把具有这一结构的动物称为半索动物。口后是咽部,在外形上相当于鳃裂区,其背侧排列着许多(7~700)成对的外鳃裂,每个外鳃裂各与一U字形内鳃裂相通,然后再由此通向体表。彼此相邻的鳃裂间布有丰富的微血管,虫体在泥沙掘进过程中,水和富含有机物质的泥沙被摄入口内,水经内鳃裂从外鳃裂排出时,就完成了气体交换的呼吸作用(图13-4)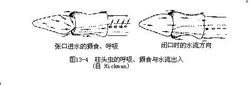 ,而食物的消化和吸收情形,则与蚯蚓大致相同。胃的分化不显著,在肠管靠后段的背侧有若干对黄、褐、绿等混合色彩的突起为肝盲囊(he-patic caecum),故称肝囊区,肝盲囊是柱头虫的主要消化腺。肠管直达虫体末端,开口于肛门。1
,而食物的消化和吸收情形,则与蚯蚓大致相同。胃的分化不显著,在肠管靠后段的背侧有若干对黄、褐、绿等混合色彩的突起为肝盲囊(he-patic caecum),故称肝囊区,肝盲囊是柱头虫的主要消化腺。肠管直达虫体末端,开口于肛门。1
3.循环和排泄:循环系统属于原始的开管系,主要由纵走于背、腹隔膜间的背血管、腹血管和血窦组成。血液循环方式与蚯蚓类似,背血管的血液向前流动,腹血管的流向往后。背血管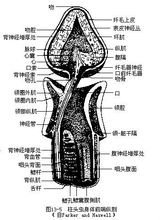 在吻腔基部略为膨大呈静脉窦,再往前则进入中央窦。中央窦内的血液通过附近的心囊搏动,注入其前方的血管球(脉球 glomer-ulus),由此过滤排出新陈代谢废物至吻腔,再从吻孔流出体外。自血管球导出4条血管,其中有2条分布到吻部,另2条为后行的动脉血管,在领部腹面两者汇合成腹血管,将血管球中的大部分血液输送到身体各部。1
在吻腔基部略为膨大呈静脉窦,再往前则进入中央窦。中央窦内的血液通过附近的心囊搏动,注入其前方的血管球(脉球 glomer-ulus),由此过滤排出新陈代谢废物至吻腔,再从吻孔流出体外。自血管球导出4条血管,其中有2条分布到吻部,另2条为后行的动脉血管,在领部腹面两者汇合成腹血管,将血管球中的大部分血液输送到身体各部。1
4.神经:除身体表皮基部满布神经感觉细胞外,还有2条紧连表皮的神经索,即沿着背中线的一条背神经索和沿着腹中线的一条腹神经索2。背、腹神经索在领部相联成环。背神经索在伸入领部处出现有狭窄的空隙,由此发出的神经纤维聚集成丛,这种结构曾被认为是雏形的背神经管,该特点表明它们似与更高等的脊索动物具有一定亲缘关系(图13-5)。1
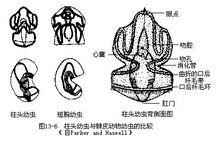 5.生殖和发育:雌雄异体。生殖腺的外形相似,均呈小囊状,成对地排列于躯干前半部至肝囊区之间的背侧。性成熟时卵巢呈现灰褐色,精巢呈黄色。体外受精,卵和精子由鳃裂外侧的生殖孔排至海水中。柱头虫的卵小,卵黄含量也少,受精卵为均等全裂,胚体先发育成柱头幼虫(tornaria),然后经变态为柱头虫。柱头幼虫体小而透明,体表布有粗细不等的纤毛带,营自由游泳生活,它们不论在形态或生活习性方面均酷似棘皮动物海参的短腕幼虫(bipinnaria)(图13-6)。
5.生殖和发育:雌雄异体。生殖腺的外形相似,均呈小囊状,成对地排列于躯干前半部至肝囊区之间的背侧。性成熟时卵巢呈现灰褐色,精巢呈黄色。体外受精,卵和精子由鳃裂外侧的生殖孔排至海水中。柱头虫的卵小,卵黄含量也少,受精卵为均等全裂,胚体先发育成柱头幼虫(tornaria),然后经变态为柱头虫。柱头幼虫体小而透明,体表布有粗细不等的纤毛带,营自由游泳生活,它们不论在形态或生活习性方面均酷似棘皮动物海参的短腕幼虫(bipinnaria)(图13-6)。
 变态时期,幼虫沉至海底,身体逐渐转为黄色,纤毛带也相继消失,前后两端分别延伸成吻部和躯干部,最终发育成柱头虫。美国沿海的有些种类(纤吻柱头虫Saccoglossus)于胚胎发育过程中不经幼虫时期和变态,即可直接发育为柱头虫。
变态时期,幼虫沉至海底,身体逐渐转为黄色,纤毛带也相继消失,前后两端分别延伸成吻部和躯干部,最终发育成柱头虫。美国沿海的有些种类(纤吻柱头虫Saccoglossus)于胚胎发育过程中不经幼虫时期和变态,即可直接发育为柱头虫。
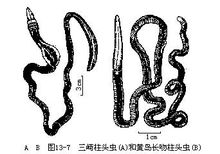
我国北部沿海分布的柱头虫有殖翼柱头虫科(Ptyroboderidae)的三崎柱头虫及玉钩虫科(Harrimaniidae)的黄岛长吻柱头虫(Dolichoglossus hwangtauensis)等(图13-7)。1
羽鳃纲头盘虫,杆壁虫,体形小,有腕和触手,营海底固着的群体生活。
已知羽鳃纲约有20种,大多是固着于深海海底营群居的小型半索动物,体长2mm~14mm,代表动物有头盘虫(Cephalodiscus dodecalophus)和杆壁虫(Rhabdopleura)等(图13-8)。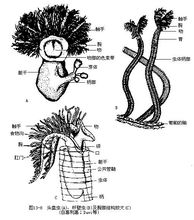 头盘虫的体长仅2mm~3mm,群栖于一个有许多小孔的公共管鞘内,彼此营独立生活,即使由出芽生殖的芽体,在成长后从亲体上脱落也能自立生活。头盘虫的身体分吻、领、躯干三部,因吻形扁平似盘而得名。该虫与柱头虫的主要不同处是领背部有腕状突起,腕上附生5~9对羽状触手,可由此摆动造成水流,使食物沿着腕和触手上的食物沟导入口内;领部只有大神经节以及分布到吻部和腕状突起的神经,但神经并不形成管状,也无空隙;咽部有一对鳃裂;肛门因肠道弯向背面呈U形而在领背开口。雌雄异体,有生殖导管输送卵和精子至体外受精,除有性生殖外,也以出芽方式进行无性繁殖。1
头盘虫的体长仅2mm~3mm,群栖于一个有许多小孔的公共管鞘内,彼此营独立生活,即使由出芽生殖的芽体,在成长后从亲体上脱落也能自立生活。头盘虫的身体分吻、领、躯干三部,因吻形扁平似盘而得名。该虫与柱头虫的主要不同处是领背部有腕状突起,腕上附生5~9对羽状触手,可由此摆动造成水流,使食物沿着腕和触手上的食物沟导入口内;领部只有大神经节以及分布到吻部和腕状突起的神经,但神经并不形成管状,也无空隙;咽部有一对鳃裂;肛门因肠道弯向背面呈U形而在领背开口。雌雄异体,有生殖导管输送卵和精子至体外受精,除有性生殖外,也以出芽方式进行无性繁殖。1
杆壁虫是群居于角质管鞘内的羽鳃纲动物,其主要特征是虫体以柄彼此相连;领背部仅有腕状突起一对;无鳃裂。
我国海域中至今尚未发现本纲动物。1
笔石纲3
笔石动物,个体小,是一类已经绝灭的海生群体动物,是奥陶纪最奇特的海洋动物类群,生活于中寒武纪到早石炭纪。有着几丁质硬壳,经石化升馏作用而保存为碳质薄膜化石。
生活习性全部海产,营底栖生活。
在动物界的位置半索动物在动物界究竟处在什么地位?这个问题直到现在还是有争论的。
半索动物除腹神经索外,还具有背神经索,并且在其前端出现空腔,呈管状,曾经被认为这是早期背神经管的雏形;它们呼吸器官为咽鳃裂;由口腔背面向前伸出一条较短的盲管,称“口索” ,过去曾被认为是原始的脊索,现在研究证明,它与脊椎动物的脊索既不同功,又不同源,可能是一种内分泌器官。4
因此把口索直接看成是与脊索相当的构造,还欠说服力,因为根据一些研究报告,口索很可能是一种内分泌器官。在另一方面,半索动物却具有一些非脊索动物的结构,例如腹神经索、开管式循环、肛门位于身体末端等等。1
半索动物门栖息于浅海和潮间带,身体蠕虫状,多穴居在泥沙中,体长2.3毫米~2.5米,已知90余种。包括体呈蠕虫状的肠鳃纲Enteropneusta和形似苔藓动物的羽鳃纲Pterobranchia两大类,77%以上属前者,最常见的就是各种柱头虫。4
就已有的研究资料来看,把半索类作为脊索动物中的一个类群,不如把它作为无脊索动物中的一个独立的门较为合适。现有的动物学文献表明:半索类和棘皮动物的亲缘更近,它们可能是由一类共同的原始祖先分支进化而成。根据是:
1.半索动物和棘皮动物都是后口动物。
2.两者的中胚层都是由原肠凸出形成。
3.柱头虫的幼体(柱头幼虫)与棘皮动物的幼体(例如短腕幼虫)形态结构非常相似。
4.有人认为,脊索动物肌肉中的磷肌酸(phosphagen)含有肌酸[creatine,NH:C(NH2)N(CH3)(CH2CO2H)]的化合物,非脊索动物肌肉中的磷肌酸含有精氨酸[(arginine,H2NC(∶NH)NH(CH2)3·CH(NH2)CO2H)]的化合物。但海胆和柱头虫的肌肉中都同时含有肌酸和精氨酸。说这两类动物有较近的亲缘关系,从生化方面也可以得到证明。
半索动物门的两个纲,在外形上差别很大。肠鳃纲的动物像蚯蚓,羽鳃纲的动物像苔藓虫。这是因为它们各自适应不同的生活环境而产生的结果,凡是分类地位很近的动物,由于分别适应各种生活环境,经长期演变终于在形态结构上造成明显差异的现象,特称为适应辐射(adaptiveradiation)。在动物界,这样的例子很多。1
研究价值在进化和生态学上有重要的研究价值,有助于了解物种的起源与发生,有助于认识生物圈与生态系统。
本词条内容贡献者为:
刘勇 - 副教授 - 西南大学资源环境学院
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国