

介形虫属甲壳纲,可以生活在淡水、微咸水和正常海水中,大部分为营底栖生活。底栖介形虫的壳由含有多于5%的MgCO,的富几丁质的方解石组成,浮游介形虫的壳为有机质,因此浮游介形虫的壳很难保存下来。1
简介
英文正式名称:ostracod或ostracode
介形虫纲(Class Ostracoda)的甲壳动物,分布广。有似软体动物的两瓣外壳。躯干极短,外表不分节,这与多数甲壳动物不同。几乎整个身体包在外壳内。据估计,现生种数量大约在1万到1.5万之间,包括海生、淡水生和陆生种类。已知的灭绝的和现生的介形虫共有65,000种,从早奥陶世到近代已发现许多化石。由于其数量多,地理分布广,因此在地层中留下丰富的化石,可作为地层的指示化石。有的属通常用来指示地下石油的勘探,与海洋地层中有孔虫的情形有些类似。现生介形虫的成虫约0.3-5毫米长,太平洋最大的大介虫(Gigantocypris agassizi)长23毫米(约1寸)。在海洋中,多数种生活在近海底处。在陆地上的湖泊、沼泽、池塘、河流、甚至土壤间隙水(例如水井)中都有分布。有的吃微生物和有机碎屑;有的吃小型无脊椎动物;少数寄生。多数喜欢在浅水中,但有的采自2,000公尺(约6,600尺)深处。
一般特征节肢动物门甲壳动物亚门介形虫纲,个体微小(多0.4-2mm)。软体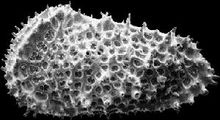 被包在两瓣壳内。壳形多样。躯体两侧对称,不分节,被包在两个壳瓣之间。壳面光滑或具各种瘤、槽、刺、脊以及其它纹饰,但不具生长线(这是与软体动物外壳的区别)。 器官发育较完善,身体分头胸部和腹部。具附肢, 末端具尾叉。两壳瓣可不相等,壳质分两层外层--钙质层;内层--几丁质,边缘可钙化。有的种具雌雄双形现象。生活领域广泛,能适应淡水、海水等多种生活环境。自早奥陶世出现,一直到现代还相当繁盛。(也有观点认为出现于寒武纪。争论的焦点在于高肌介类是否属于介形虫。)
被包在两瓣壳内。壳形多样。躯体两侧对称,不分节,被包在两个壳瓣之间。壳面光滑或具各种瘤、槽、刺、脊以及其它纹饰,但不具生长线(这是与软体动物外壳的区别)。 器官发育较完善,身体分头胸部和腹部。具附肢, 末端具尾叉。两壳瓣可不相等,壳质分两层外层--钙质层;内层--几丁质,边缘可钙化。有的种具雌雄双形现象。生活领域广泛,能适应淡水、海水等多种生活环境。自早奥陶世出现,一直到现代还相当繁盛。(也有观点认为出现于寒武纪。争论的焦点在于高肌介类是否属于介形虫。)
现代介形虫主要构造
软体介形虫身体两侧对称,不分节,外被两瓣壳包裹,个体虽小,但结构复杂,各种器官发育比较完善,身体分为头胸部和腹部,二者的接合处不明显。头部大,约占身体大小的一半,胸部末端生有一对 尾叉。眼位于头部背侧,通常为单个的眼,多数介形虫不具眼,有的属种幼年期具眼,到成年期消失。介形虫身体某些部位生有感觉毛,通过壳壁的毛细血管伸出壳外司感觉作用。消化系统位于头部腹侧的口、食道、胃、肠及位于身体后端的肛门组成。无腮,通过薄的体壁自然扩散进行呼吸。除了个体较大,浮游类型的目之外,均缺少血管和心脏。肌肉系统复杂。
尾叉。眼位于头部背侧,通常为单个的眼,多数介形虫不具眼,有的属种幼年期具眼,到成年期消失。介形虫身体某些部位生有感觉毛,通过壳壁的毛细血管伸出壳外司感觉作用。消化系统位于头部腹侧的口、食道、胃、肠及位于身体后端的肛门组成。无腮,通过薄的体壁自然扩散进行呼吸。除了个体较大,浮游类型的目之外,均缺少血管和心脏。肌肉系统复杂。
头和胸部具7对分节的附肢,除第一触角外,均为双枝型,包括两个明显的分支:外侧的外肢和内侧的内肢。附肢上具刚毛,末端为爪。头部生有4对附肢,位于前面的第一触角,为长而向前逐渐变细的单肢型附肢,用于爬行、游泳和摄食。其后第二触角用于爬行、游泳和攀登。口附近的双肢型大颚基部坚硬具强齿,帮助咀嚼食物。口后边双肢型小颚可以搅动水体提供滤食水流,同时加强动物体周围的水循环有助于呼吸作用。胸部生有3对附肢,皆为双肢 型起源、但外肢退化,主要用于爬行、挖掘、进食以及交配。
型起源、但外肢退化,主要用于爬行、挖掘、进食以及交配。
繁殖和个体发育雌雄异体。有的种类无雄性,以单性生殖方式产生后代,有些属种行两性生殖,也有两性生殖和单性生殖都可以进行。雌性生殖器官由位于身体后部的卵巢及输卵管组成。在特定季节生殖。卵呈圆形或卵圆形,耐干旱及寒冷性很强,利于适应低温、干旱环境,并可较长距离扩散,在适宜的温度下孵出幼虫。它的增长是不连续的由卵孵化出幼虫称第一龄期,当身体生长对壳体来说已经过大时,坚硬的双壳脱落,随之身体迅速增长和发育,同时形成新的壳并硬化。由第一期至成年,通常经过8个这样的龄期,蜕壳8次,每蜕壳一次,个体增大一些,各器官、附肢不断增加、完善,最后达到成年。期发育过程中,壳形、壳饰及各种构造特征也发生一定规律性的明显变化。
壳根据运动前进方向分左、右两瓣
壳的成分与构造介形虫的壳主要成分为钙质和几丁质。碳酸钙为主要成分,占95%以上。不同地史时期壳的各种元素含量变化较大。可分为外壁和内壁。壳可分为 上表皮、外表皮、内表皮和表皮细胞四层。上表皮极薄,不能保存为化石;外表皮为钙质。常保存为化石;内表皮也为钙质;表皮即由节肢动物的表皮细胞组成。内壳层由几丁质组成,甚薄,仅边缘部分钙化成较厚的钙化擘,起着增进壳的自由边缘闭合的作用。幼年期和某些淡水介形虫及浮游的丽足介目的大多数种属的壳质钙化弱,不一保存为化石。
上表皮、外表皮、内表皮和表皮细胞四层。上表皮极薄,不能保存为化石;外表皮为钙质。常保存为化石;内表皮也为钙质;表皮即由节肢动物的表皮细胞组成。内壳层由几丁质组成,甚薄,仅边缘部分钙化成较厚的钙化擘,起着增进壳的自由边缘闭合的作用。幼年期和某些淡水介形虫及浮游的丽足介目的大多数种属的壳质钙化弱,不一保存为化石。
壳的外形、壳饰及双形现象侧视有近圆形、半圆形、椭圆形、肾形、纺锤形、三角形、菱形及梯形等。背视主要为各种凸度的透镜体及椭圆形等。端视多为椭圆形和三角形。多数两壳不等大,较大的壳常以边缘叠覆与较小的壳上,称为超覆或叠覆。左壳边缘叠覆与右壳称为左超覆,反之,右超覆。某些壳面光滑,多数壳面 具各种不同纹饰。位于前背部的圆形小结节或小坑称眼痕。具双形现象,即同一种的雌雄不同壳体的形态特征往往存在显著差别。
具各种不同纹饰。位于前背部的圆形小结节或小坑称眼痕。具双形现象,即同一种的雌雄不同壳体的形态特征往往存在显著差别。
铰合、肌痕、边缘构造壳内侧构造:铰合构造--壳内侧、背部边缘、突起的齿、脊和凹入的齿窝、槽组成
肌肉印痕--闭壳肌痕、大额肌痕
边缘构造--内缘板、结合带、结合线、毛细管带
这些都具有重要的鉴定意义
系统分类分类节肢动物门(Arthropoda) 甲壳超纲(Crustacea)
甲壳超纲(Crustacea)
介形虫纲(Ostracoda)
高肌介目(Bradoriida,目前认为不是介形虫)
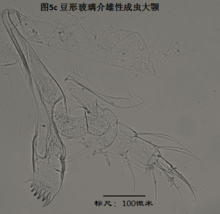
豆石介目(Leperditicopida,目前认为不是介形虫)
古足介目(Palaeocopida)
速足介目(Podocopida)
丽足介目(Myodocopida)
介壳化石的分类依据1、介壳形状
2、壳面装饰
3、肌肉印痕
4、两壳的叠覆关系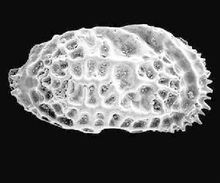
5、铰合构造(很少用)
6、内边缘构造
7、眼点
地史分布介形虫的近亲之一,高肌介类从寒武纪早期出现,化石分布于亚、欧、澳及北美地区。但目前的主流观点认为高肌介不属于介形虫。目前已知的最早出现的介形虫,发现于距今4.85亿年前的早奥陶世地层中的古足介类。奥陶纪是古足介目全盛时期。三叠纪时速足介目开始占统治地位。侏罗纪时,浪花介超科、金星介超科、小浪花介科等大量繁盛。同时,达尔文介超科、土菱介超科等仍继续发展。白垩纪开始,介形虫动物群再次出现大发展时期。更新世至现代介形虫属的数量巨大。
生物地层意义:中新生代海陆相地层中介形虫可分为很多组合,具有重要的地层学意义。
生态与应用湖相介形虫古生态学应用1、研究介形虫种属的分类、组合、丰度及分异度、壳体的大小、形态、结构、壳饰及厚度;
2、分析水化学条件: 盐度、离子组成、温度、深度、食物的供给等;
3、水体的温度、盐度、酸碱度影响介形虫生态特征;
4、介形虫的古生态学,可以提供重要的环境变化信息,掌握盐度和离子组成对介形虫种属变化的影响;
5、了解介形虫生命历史和生态消长的过程。
海洋与湖泊的测深仪要想测量海洋的深浅,除了使用各种测深仪实际测量外,还可从介形虫得到大海深浅的数据。介形虫有自己固定的栖息地,从不到处漂泊,例如在深海生活的种类决不到浅海处栖居;在浅海处生活的种类也决不到深海去邀游。地质学家根据介形虫的这一习性,就能估算出大海的深浅。例如,在我国南黄海西北部地区海底泥沙中介形虫的分布,南部以中华丽花介为主,北部以穆赛介为主,东部以克利特介为主。这三种介形虫分别生活在0-20m、20-50m和大于50m水深的海区。因此,根据这些介形虫种类的分布情况,就能绘制出一幅简单的海底地形图。
对于现代的海洋测量来说,介形虫给出的深浅数据当然是太粗略了,根本无法与现代仪器的精密测量相提并论。但是,介形虫具有测量千百万年前海水深浅的本领,却是任何现代精密测深仪都望尘莫及的。现代测深仪无论多么先进,只能测现代海洋的深浅,对于遥远地质年代的海洋的深浅则无能为力。在漫长的历史进程中,海洋早已发生了巨大的变化。面对这个面目全非的海洋,介形虫却能大显身手。比如,地质学家从地中海几千万年前形成的沉积物中,发现了一种叫深海角介的只能在大洋里生活的介形虫,而在年代更新的沉积物中,却再也见不到它的踪迹。由此得知,古地中海曾经是一个大海,与大西洋相通,水深可能达到几千米,以后它又与大洋失去联系,封闭成如今名副其实的被陆地包围着的地中海。在这一点上,介形虫所提供的宝贵数据是无与伦比的。
考古发现2014年5月14日,外媒报道,澳大利亚昆士兰州发现了世界上最古老且保存得最完好的精子,该精子藏身于距今约1700万年的化石里。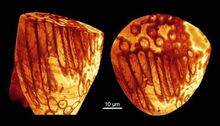
据悉,这一精子是在一只已经石化了的雌性远古物种介形虫的性器官内发现的,科学家称这只介形虫在死前刚刚交配过。相对于这种动物的体型而言,该精子显得非常大。
精子藏身的化石由三名研究人员于数年前发掘。其中一名研究员阿彻教授(Michael Archer)表示,99.9%微生物的精子与体积相比都非常小,而且这只介形虫的软组织能保存得如此完好,令我们感到异常惊讶。
“当捡起这些坚硬的、干燥的石头时,我们通常会用锤子敲它。我们完全没有想过在这些石头里会发现曾存在于软组织内的精子。”阿彻教授说。
阿彻教授亦指出,目前尚不清楚这只介形虫的细胞是如何变成化石的。但蝙蝠分泌出来的液体在保存精子的过程中可能发挥了重大作用。而蝙蝠液体大量存在于发现化石的洞穴地面。
关于1. 简介
介形虫是一类小型甲壳动物,属于节肢动物门、甲壳动物亚门、介形虫纲。它们的整个身体包被在一对外壳中。外壳的长度一般介于0.3–5 mm之间,主要成分通常是方解石和几丁质。身体由头、胸、腹部构成,但体表不可见分节。通常具8对附肢,除第一触角外,均为双枝型起源。现生介形虫广泛分布于各种水域。目前发现的最早的介形虫化石属于4.85亿年前的奥陶纪。
注:1)由拉丁文Ostracoda派生出介形虫的两个英文词:ostracod是除法国以外的欧洲、以及澳洲的写法,ostracode是法国及北美的写法。拉丁文Ostracoda的首字母须大写,是分类学的用法,表示介形虫纲。
2)过去,人们为甲壳动物设甲壳纲,为介形虫设亚纲。近年始变。
3)介形虫的附肢自前而后有:第一触角、第二触角、大颚、小颚、第一胸肢、第二胸肢、第三胸肢、尾肢。由于体表不可见分节,因此所谓的第一胸肢属胸部还是头部尚有争议。
4)介形虫的第一触角是单枝型的,其余触角是双枝型的,尽管双枝型的部分特征有时退化至不可见。这与其它甲壳动物一致。双枝型附肢的基本构造包括原肢(protopod)以及由原肢分出的外肢(exopod)和内肢(endopod)。此外,原肢上有时还着生有内叶(endite)和上肢(epipod)等结构。
5)尾肢(uropod)曾被称为furca,并被认为起源于身体的尾节(telson)。但现在认为它起源于腹部一对附肢的振动片(vibratory plate)(Meisch, 2007)。它也是多数介形虫唯一的腹部附肢。在某些浪花介超科的雄性中,尾肢之前还有一对称为brush organ的附肢。
6)上面所说的8对附肢不包括由附肢演化而来的阴茎等结构。
7)目前公认的最早的介形虫是出现于奥陶纪早奥陶世的古足介类(Palaeocopida)。出现于寒武纪的高肌介类(Bradoriida)不被视作介形虫。详见下文。
2. 分类系统
如图1所示,介形虫纲下分四个目:古足介目Order Palaeocopida Henningsmoen 1953,丽足介目Order Myodocopida Sars 1866,平足介目Order Platycopida Sars 1866,以及速足介目Order Podocopida Sars 1866。高肌介目Bradoriida和豆石介目Leperditicopida。是否应划入介形虫纲,还存在较大的争议,原因就在于,这两类动物都已经灭绝,留下的化石大多是外壳,很难找到保存良好的软体,从而根据附肢的解剖结构来判断其分类地位。从目前发现的唯一的高肌介软体化石、来自云南澄江寒武系的Kunmingella douvillei(Hou et al., 1996)来看,其附肢结构不同于普通介形类(Williams et al., 2008)。因此,目前的主流学派把高肌介排除在介形虫纲之外。至于豆石介,笔者不清楚是否有软体发现,但介形虫学界似乎也倾向于不将它纳入介形虫。
上面的这个争议也引发了另外一个问题,即最早的介形虫到底是出现于寒武纪还是奥陶纪。因为高肌介出现在寒武纪,而目前公认的属于介形虫的四个目中,最早出现的是古足介目——发现于距今4.85亿年的早奥陶世地层中(Williams et al., 2008)。如果高肌介属于介形虫,那么就应该说,最早的介形虫出现于寒武纪;如果高肌介不是介形虫,那么介形虫则应起源于奥陶纪。由于一般认为高肌介不属于介形虫,所以,一般也认为,介形虫起源于奥陶纪、而不是寒武纪。
但是,值得再次强调的是,高肌介和豆石介的分类地位是悬而未决的问题。以高肌介目前仅有的附肢化石证据,即Kunmingella douvillei化石,尚不足以将所有的高肌介排除在介形类以外。随着新化石的发现,将来很有可能在地层中发现属于介形类的其它的高肌介软体化石。分子生物学证据也暗示,介形类的起源很可能比目前发现的4.85亿年要早得多(Regier et al., 2005)。因此,目前所发现的4.85亿年前的古足介,很可能不是最早的介形虫;更早的介形虫可能另有其“虫”。
根据Ostracon社区里介形虫学家的推测,目前地球上生活着大约10000到15000种现生介形虫,其中大约三分之二在海洋里,三分之一在各种陆地水域中。在现生介形虫的三个目中,速足介最为繁盛。目前我们在各种陆地水域中发现的介形虫,几乎无一例外地属于速足介。速足介目又分为三个超科(图1):达尔文介超科Superfamily Darwinuloidea Brady & Norman 1889,金星介超科Superfamily Cypridoidea Baird 1845和浪花介超科Superfamily Cytheroidea Baird 1850。这三个超科中又以金星介种类最多——至少在陆地水域中是这样。
在金星介超科中,玻璃介科Family Candonidae Kaufmann 1900和金星介科Family Cyprididae Baird 1845是大科。就非海洋种类来说,玻璃介科已知的种类有大约550种,金星介科有大约1000种,占所有三个超科非海洋介形虫种类的75%(Martens et al., 2008)。

 3. 软体形态结构
3. 软体形态结构
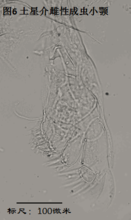
介形虫可以分为外壳和软体两部分。外壳主要是方解石,可在地层中保留成为化石,为古生物学家、地层学家、古气候学家所感兴趣;软体分为身体和附肢,以及由附肢所演化而形成的阴茎等器官,是现代生物学家研究的对象。现生介形虫分类的主要依据是其成虫附肢的形态结构,而化石介形虫的分类,则几乎只能依据外壳。依据外壳进行的属种鉴定有一定的不确定性,尤其是对于玻璃介科、土星介属等类群。下面以金星介科异星介属、土星介科土星介属、以及玻璃介科豆形玻璃介属为例,简要介绍介形虫的软体结构。所有标本均为笔者采自国内的成虫,所有照片均为笔者拍摄。下文描述的结构模式,除加以说明外,仅适用于金星介科。
1)第一触角,antennule (A1),见图3
分为8节(图3)。第一和第二节融合,因此有人又把这两节合称第一节,而把第一触角分为7节。与其它甲壳动物一样,介形虫的第一触角在起源上可能不同于其它附肢,因为它是单枝的(uniramous),没有内肢(endopod)和外肢(exopod)之分;而其余附肢是双肢(biramous)的,有内肢和外肢之分。
大多数金星介超科种类在每节的刚毛数目和位置上都是一致的。但其中某些豆形玻璃介的第一触角刚毛式则有所不同:它在第一节上多出一根背毛,第三节、第四节、第五节、第六节各少一根腹毛,第七节也少一根毛。另外,至于每根毛的长短、粗细,以及其上是否着生小毛,也因属种而异。
第一触角的刚毛式在金星介科的不同介形虫之间被认为是比较保守的,因此很少有人用它来对介形虫进行分类。但实际上,某些毛的长度或可用作介形虫分类的依据。
第一触角主要用于运动(游泳、钻掘等)和感知。
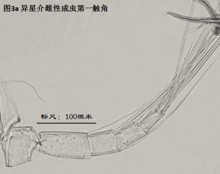
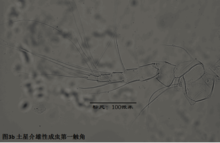
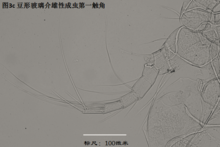
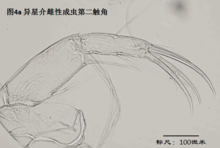
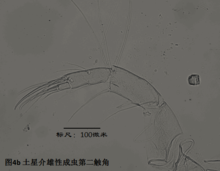
2)第二触角,antenna (A2),见图4
与其它甲壳动物一样,介形虫的第二触角是双枝的,有内、外肢之分,它们是着生在原肢(protopod)上的两个分枝,内肢靠内,外肢靠外。其中外肢退化为一长两短三根刚毛,而内肢则粗壮,构成附肢的主干。
第二触角共有6节,其中原肢有2节、相互融合,内肢有3-4节。许多种类的第二触角形态存在显著性别分化。第二触角上着生的大多数刚毛和爪子(爪子可认为由刚毛演化而来)比较保守,但第3节末端内侧着生的6根游泳刚毛的长度,第4a节末端内侧着生的4根t系列刚毛,以及第4b和5节末端着生的z系列的刚毛(某些种类雄性的z1和z2刚毛变为爪子)和G系列的爪子(某些种类雄性的G3细小,呈刚毛状)的形态,是较为重要的分类学特征。

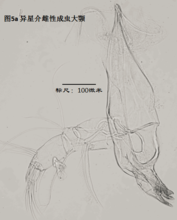
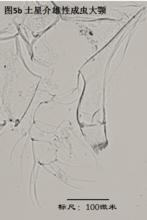
3)大颚,mandible (Md),见图5
大颚由coxa(masticatory process),振动片(vibratory plate)以及palp组成。其中coxa相当于原肢,vibratory plate由外肢演化而来(Karanovic, 2012),palp则相当于内肢。通常认为,内肢的刚毛式是重要的分类学依据,尤其是其S系列的刚毛(着生于内肢第一节),以及α,β,γ(分别着生在内肢的第一、第二和第三节)刚毛的形态。但实际上大颚的刚毛很多,常重叠在一起,观察起来很费劲。
大颚用于摄食、咀嚼,有的种的大颚也用于运动。
4)小颚,maxillule (Mxl),见图6
基本构成与大颚相似,由endite(可能相当于大颚coxa上的牙齿),称为palp、分2节或不分节的内肢,以及由外肢演化而来的振动片(Horne, 2005)组成。
小颚用于摄食、咀嚼。其振动片较为发育,用于划动水流、从而使机体获得足够的氧气。
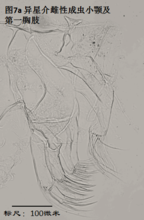
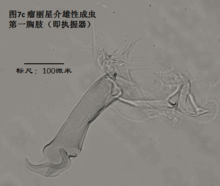
5)第一胸肢,the first thoracopod (T1或L5),见图7
基本构成与前两对附肢相似。由masticatory process(可能相当于endite),分节或不分节的palp,以及振动片组成。endite位于前方,palp和振动片位于后方。
雌性第一胸肢没什么可说的。用于摄食(浪花介超科的用于行走)。
雄性第一胸肢的palp,在许多金星介超科中特化为执握器(clasping organ,或prehensile palp),常较为粗壮,用于在交配中抓住雌性。左右执握器常不对称。由于执握器是与生殖相关的器官,因此不同种的执握器常差异明显。有经验的介形虫学家有时仅仅根据执握器就能鉴定介形虫的种。
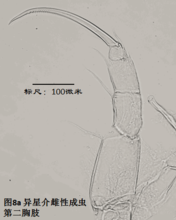
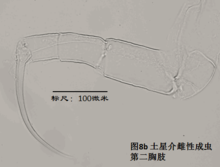
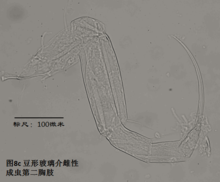
6)第二胸肢,the second thoracopod (T2或L6),见图8
在所有速足目介形虫中,这对附肢都是用来走路的,称为步足(the walking leg)。共有6节。原肢实际上是2节融合在一起,每节的前侧的末端分别着生一根毛,分别称为d1和d2。笔者不清楚d2是否是外肢。内肢共有4节,但有时第二、第三节内肢(即第二胸肢的第四、第五节)相互融合。这种情况与第二触角有些类似;但融合并非由于性别的缘故,而是在某些种的雌雄都如此,例如隆起土星介(Ilyocypris gibba)。内肢的每一节,不论它与相邻的节融合与否,在其前侧的末端都着生有一根有名字的毛:e,f,g和h1。倒数第二节,除了g外,还有一根没有名字的毛。在最末一节,除h1外,还着生有一根粗壮的爪子h2和后侧的毛h3。最末一节通常为一爪两毛,偶尔是两爪一毛。
步足的刚毛数目、位置和长短是重要的分类学依据。
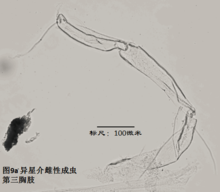
7)第三胸肢,the third thoracopod (T3或L7),见图9
其结构与第二胸肢明显是同源的,但原肢末端多了一根毛,dp。金星介超科的第三胸肢向后、向上蜷曲,特化为一对清洁足(the cleaning leg),用于清洁介壳内部。两节原肢的融合线常不可见。与第二胸肢类似,第三胸肢的时第二、第三节内肢(即第四、第五节)常完全融合以至于融合线不可见。在金星介科中,第四五六节完全融合,并且g和h2刚毛特化,h1很短,末端形成一个“pincer organ”(图9a)。
注意:由于步足和清洁足的融合很完全,通常无痕迹,并且在同一种的两性间是一致的,因此习惯上把融合的节合称一节;这与第二触角4a和4b即使融合仍分开标注的习惯不同。(由于第二触角仅在某些种的雄性中有分界线,雌性中都是融合的,因此如果不分开标注,就会造成雌雄个体对比描述上的混乱。)
清洁足的刚毛数目、位置和长短也是重要的分类学依据。
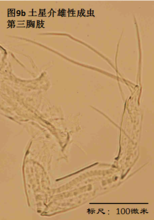
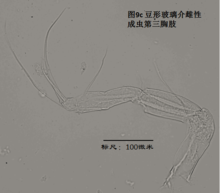
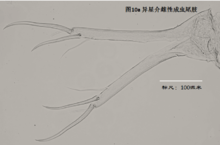
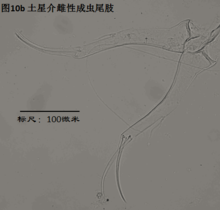
8)尾肢,uropod (Ur),见图10
由一根不分节的主干(uropodal ramus,复数rami),两个爪Ga和Gp,以及两根毛Sa和Sp组成。a意为anterior,前方,顾名思义,Ga和Sa是位于前面的;p意为posterior,后方,也就是说Gp和Sp是位于后侧的。
ps: 介形虫的尾肢通常向下悬着,并斜指前下方。
尾肢主要用于运动(游泳或行走)。
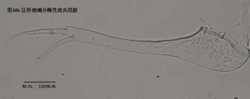
9)生殖器官。
雌、雄的外生殖器都成对出现,并且都位于第三胸肢和尾肢之间。
雄性阴茎,penis,更严格的提法是hemipenis (Hp),指一对阴茎中的一个,见图11
与我们通常所认识的“阴茎”的概念有所不同,介形虫的阴茎不光包括用于插入雌性外生殖器的部分,还包括很多其它的结构。但这些结构的作用知之尚少(Karanovic, 2012)。
阴茎在不同的介形虫之间差异甚大。即使在同一个属的不同种之间,阴茎也有明显差别。这种差异远大于其它附肢的差异。这是因为,阴茎是用于交配的,而不同物种必须不能互相交配、具有生殖隔离。这种隔离机制主要是通过交配(包括前戏,即同一物种不同性别的识别,mate recognition)过程中同一物种雌雄个体能够互相配合、完成交配,而不同物种之间不能来实现。一个简单的例子,试想,阴茎的一部分要插入雌性外生殖器,因此其形状、大小和所着生的位置,就要恰好与雌性阴道吻合。这种关系就好比“钥匙和锁”的关系。锁不同,钥匙也不同。
由于不同种的阴茎差别很大,因此它是十分重要的分类学特征。常常仅根据阴茎就能轻松鉴定介形虫。但是有些介形虫类群只有雌性,进行孤雌生殖,因此只有通过对其它附肢的详细观察来定种。
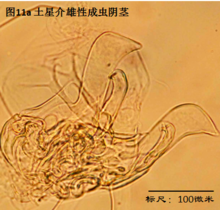
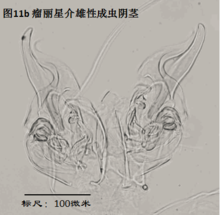
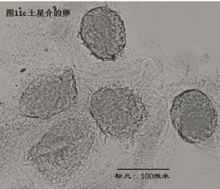
4. 生态
介形虫广泛分布于海洋、湖泊、池塘、沼泽、溪流、地下间隙水等各类水域中,极少数种类为陆生或半陆生。多为底栖种类,在底泥表面爬行、钻入底泥浅层,或攀爬于水草之上。许多种类在第一和第二触角上生有长的游泳毛,能游动,但活动范围仍多限于水底层附近,绝少来到水的中上层。因此,用浮游生物网很难捕到介形虫。短背介这一类除外;它们介壳的腹部是平的,能倒悬并紧贴于水面之下。
食性杂,取食各种有机碎屑。但也许不同种或多或少有食性偏好吧。
有些种类可寄生于两栖类和鱼类;有寄生在人类肛门的报道。
对水温、水的化学成分(离子组成、含氧量等)都有一定要求,不同属种的适应范围也不一样。
5. 研究历史
早在1748年,生物分类学的奠基人、瑞典自然学者Carolus Linnaeus,在Systema Naturae一书中,就对介形虫动物进行了初次描述(Tressler, 1959)。但Linnaeus未对所描述的标本进行正式命名,后人只是根据他的描述,得知该标本属于现在所称的介形虫。
1776年,丹麦人Otto Friedrich Müller描述了一些小型甲壳动物,将其归入一个新设立的属Cypris(金星介属,Genus Cypris)中,分别命名为Cypris pubera,Cypris candida(现称Candona candida)和Cypris laevis(现称Cyclocypris laevis),等等。这就是最早被正式命名的介形虫。随后,在1785年,Müller在甲壳纲下建立了Entomostraca亚纲(Subclass Entomostraca),将Cypris属和另一个新设立的属Cythere(浪花介属,Genus Cythere),以及其它一些并非介形虫的甲壳动物归入该亚纲(Baird, 1843; Hanai et al., 1985; 禹娜等,2004)。
1806年,著名的昆虫学家、被誉为“昆虫学王子”(Prince of Entomology)的法国人Pierre André Latreille,在他流放期间,建立了Ostrachoda亚纲(Subclass Ostrachoda)(也有说法认为Ostracoda亚纲是Latrielle于1802年建立的),并将介形类和枝角类从Entomostraca亚纲移入Ostrachoda亚纲。“Ostrachoda”后来被改写为“Ostracoda”,成为现在所使用的“介形虫”一词。英文的ostracod和ostracode,都是从拉丁文Ostracoda派生而来(Hanai et al., 1985)。1821年,H.E. Straus将枝角类从Ostracoda亚纲中分出,从而使Ostracoda成为独立的一类,即现在所称的“介形虫”。
继Latrielle和Straus之后,越来越多新的介形虫被发现并命名。随着资料的积累,人们对介形虫的软体和外壳结构进行更加系统和详细的描述。现生介形虫的分类系统,在十九世纪晚期和二十世纪上半叶之间逐渐完善。在二十世纪,一些学者还对介形虫的内部解剖结构和个体发育进行研究(Snodgrass, 1956)。与此同时,形态学和分类学的资料也在不断积累(Tressler, 1959)。到了二十世纪后半叶,有人提出,生物的遗传物质能记录其进化历程。由于介形虫是现存的历史最为悠久的生物类群之一,其DNA、RNA所包含的生物演化信息必然极为丰富。因此,一些学者开始通过研究细胞中的DNA、RNA来追溯介形虫及其所代表的节肢动物的演化历史和内在规律(Bode et al., 2010)。另一方面,介形虫作为广泛分布的一类小型动物,它们在生态环境测评和保护研究方面的作用也渐渐为人们所认识(Pieri et al., 2012)。
至于对介形虫化石的研究历史,则追溯到1813年。当时,“介形虫微体古生物学之父”(Father of Ostracode Micropalaeontology)、法国的Anselme Gaetan Desmarest从法国中部Allier地区的渐新世地层中发现了第一枚介形虫化石标本,命名为Cypris faba(Hanai et al., 1985; Matzke-Karasz et al., 2007)。随后,Jones(1850)在英格兰南部更新世地层中进行了最早的对第四纪介形虫化石的研究。此后,在十九世纪下半叶中,Jones,Prestwick,Sherborn,Chapman,Kirkby等人对多地的化石介形虫进行了描述,积累了大量形态学和地层学资料,由此逐步建立了化石介形虫的分类系统。
在二十世纪的大多数时间里,介形虫化石的研究着重于它们在地层中的分布、尤其是在某一个地区的地层序列中的分布,目的主要是对含化石地层进行划分对比和确定地层时代。在这一时期,由于石油地质勘查和地质矿产调查的需要,欧、美、前苏联和中国在内的世界各国,都开始利用介形虫和其它微体化石进行生物地层学的研究。在我国,介形虫生物地层学的开拓者和奠基人是侯佑堂(1919–2010)。二十世纪中后期至今,在对介形虫生物地层学深入研究的同时,古环境研究学者开始根据地层中化石属种组成和壳体地球化学的变化,推测地质时期的环境波动。
如上所述,对现生介形虫的研究和对介形虫化石的研究是有区别的。对现生介形虫的研究,主要是依靠它的“肉”,也就是介壳里的那个身体——从水里活捉介形虫,依靠附肢的形态、刚毛的数目、位置和长短等来进行鉴定和分类,并提取DNA进行研究;尽管介壳的形态也具有一定分类学价值。对化石介形虫的研究则几乎仅仅围绕介壳进行——从沉积物和地层里挖出介壳,研究不同种类的地层和时代分布,从而对地层进行对比、划分;研究特定时代介形虫的种类组成、介壳化学成分,从而推测古代环境状况。因此,现生介形虫学家和化石介形虫学家通常是不打交道的,尽管他们在某些有交叉的论题上合作。例如研究介形虫的演化时,地层中的演化谱系可以与根据现生种类亲缘关系得到的演化关系进行对比。再如,对某些现在还活着的介形虫,当它们的介壳从沉积物中被找出来的时候,对它们的属种鉴定就需要与现生种类的分类系统相互对照,统一到后者上来。Limnocythere inopinata具有不同瘤饰的各个表型,在很长时间内就曾被国内的化石介形虫学者分为不同的物种;但通过对现生种的附肢的观察发现,它们实际上是同一个东西(Yin et al., 1999)。最后,当古环境学家为地层中某种介形虫所反映的环境是怎样的而苦恼时,他们也需要请教现生介形虫学家,这个种现在生活在什么样的环境中。
就现生介形虫来说,在当前学术界,研究的中心在欧洲,而中国的研究远远落后。就化石介形虫的研究而言,我国可以说曾经一枝独秀,但或将日趋式微。
6. 应用
不同应用的罗列顺序和篇幅未必反映其重要性。
1)地层对比和划分。请见上文。
2)古环境研究。请见上文。
3)生态环境测评,以及生物多样性保护
人们利用介形虫的种类组成和物种多样性来反映水域的水质和生态健康状况(Pieri et al., 2012)。
对生态健康状况的测评,原理是这样的。在众多介形虫中,有世界广布的属种,也有地方性特有的属种。广布种适应性强,地方种通常只能适应某个地区的稳定的、原生态的环境。当某个地区的自然环境遭受破坏(通常是人为的)时,原生的生物群落就将被瓦解,而一些别的地区的物种,通常是那些适应能力强的广布种就侵入并占据了这个地方。这个过程中,生物群落的成分改变了,并且多样性通常下降,因为被破坏、被污染的环境中只有少数适应能力较强的物种能够生存。而当人们采取一些保护措施、恢复环境的水质、减少人类活动的侵扰,从而使环境逐渐好转时,这个地区的生物多样性就又会增加,一些原本濒临消失的地方性种又重获新生,并且更多来自其它地方的、适应能力并不是那么强的物种也加入到新的群落中来。人们注意到,这个次生的生物群落的组成,和人类来到之前的那个原生态的群落的组成是有所不同的,因为在破坏——恢复的过程中,某些土著物种被不可逆地淘汰掉了,而一些外来的移民则在这里扎根。尽管成分变了,生物多样性的增加仍然反映了环境保护措施的成效、生态健康状况的改善。另一方面,同样的生物多样性,那些地方性属种丰富的生境,就代表保护得很好、从未或很少遭受人类破坏的原生态状况;而那些充斥着各种广布种的生境,就很可能是曾经遭受人类活动破坏、之后逐渐修复的生境。
介形虫当然不是唯一的能用于生态测评的物种(Boets et al., 2011)。但笔者认为,介形虫、硅藻等小型生物,由于其分布广、繁殖快、数量多,因而与较大的生物相比,具有容易取样、响应灵敏、更易满足统计要求等优点。
利用生物群落对水质进行评价,是因为人们在各种水质化学指标之外,还希望看到一些直观的、利用水生生物的生存状况来反映水源好坏的指标。毕竟,新型污染物太多了,光看仪器测量有点不放心,总想看看里面的鱼虾活得怎么样。而且从心理上,现在的人们也倾向于喝那些“原生态”的水。但是利用生物来进行水质评价,取代不了器测,充其量不过是一种补充手段。
本词条内容贡献者为:
刘勇 - 副教授 - 西南大学资源环境学院
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国