

曳鳃动物已记载仅有18种,全部生活在浅海和深海的泥沙中,从潮间带到几千米深海。大部分体形较大的种类生活在冷水中;有一些小型种类(如Tubiluchus)分布较为广泛,包括热带海洋。曳鳃动物是中寒武纪海洋底栖生物(benthos)的重要成员。已描述的11个化石种,它们是寒武纪海洋中占优势的无脊椎动物。现在的种类与其寒武纪的祖先没什么大变化。 这类动物体长0.5 mm一40mm,体呈圆柱形,躯干较大,其前端有一翻吻(introvert),后端常有一或两个尾附器(caudal appendages)尾附器与体壁连续,其表面角质膜变薄,有学者认为它具有气体交换、渗透调节和化学感受的功能,但没有证据证明。神经系统:围绕咽部有一神经环和一腹神经索。在体腔中有吞噬的变形细胞、红细胞并含有蚯蚓血红蛋白(hemerythrin)。体腔液也有流体静力骨骼的作用。学者们一直认为它具有假体腔,但也有学者认为它具有真体腔,从图8—29可见它具有在中胚层之间形成的腔,但没有体腔上皮;而有假体腔的功能,因此,或者说它具有处于萌芽状态的真体腔。 曳鳃动物仅是现在开始受到关注,了解其进化关系,从其具有神经环,前端的口和翻吻以及分子证据,认为它与动吻动物、兜甲动物亲缘关系较近。1
形态特征该门动物呈蠕虫状,体长几毫米至200毫米。两侧对称,体圆柱形,不分节。 典型的曳鳃动物大多可分为吻(翻吻)、躯干部(腹部)和尾(尾附器)3个部分。吻位于虫体前端,可伸缩,口位于吻前方中央。吻表面具多排环状排列的咽齿和许多纵排的小圆锥形杯状乳突。躯干部具许多假体环,但非分节,常覆有小棘或瘤。后端腹面开口为肛门,肛门前两侧开口为泄殖孔。尾位于躯干部之后,有人认为是躯干部的延长,具中空的柄和许多囊泡状突起(尾囊),不同种的曳鳃动物尾变化较大,或仅为一细长的尾,或无尾。
典型的曳鳃动物大多可分为吻(翻吻)、躯干部(腹部)和尾(尾附器)3个部分。吻位于虫体前端,可伸缩,口位于吻前方中央。吻表面具多排环状排列的咽齿和许多纵排的小圆锥形杯状乳突。躯干部具许多假体环,但非分节,常覆有小棘或瘤。后端腹面开口为肛门,肛门前两侧开口为泄殖孔。尾位于躯干部之后,有人认为是躯干部的延长,具中空的柄和许多囊泡状突起(尾囊),不同种的曳鳃动物尾变化较大,或仅为一细长的尾,或无尾。
体壁由周期性蜕皮的角质膜、单层表皮、环肌和纵肌以及一层不清楚的膜组成。消化系统为一直管,由口、口腔、肌肉质的咽球、食道、肠、直肠、肛门等器官组成。神经系统简单,不与表皮分离,亦无神经节的分化,具围咽神经环和腹中部的神经素。排泄器官原肾型,1对,位于体后肠两侧的体腔内,具许多焰细胞,排泄管下行与生殖管相遇为泄殖管,并以泄殖孔开口于体外。
体腔大,位于肠管与体壁之间,并延伸至尾部,内充满乳白色液体,具两种游离的细胞:红细胞(内含具呼吸色素的蚓血红蛋白,可能具携氧和储氧的功能)和游动的变形细胞。
生长繁殖雌雄异体,体外受精。胚胎发育过程尚不清楚。后期幼虫与成体相似,具翻吻, 但覆盖着多片厚的角质兜甲,变态后兜甲消失。
但覆盖着多片厚的角质兜甲,变态后兜甲消失。
生活环境曳鳃动物是海洋底栖性的动物,大部分生活在靠近两极地区的冷海中,从浅海到 深海都有,自由生活,主要栖于含泥的沉积物中。
深海都有,自由生活,主要栖于含泥的沉积物中。
生活习性在无氧的泥滩、超咸水或半咸水水域等沿岸环境中可能是重要的动物组成成分。在该生境中生活可能避免与其他后生动物竞争。以翻吻在泥内吞咽小型多毛类环虫、甲壳动物和其他曳鳃动物。
亚种种类曳鳃动物门是很小的一门,已知有2纲4科12种。中国仅发现2属2种:尾曳鳃虫发现于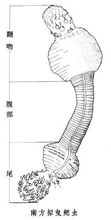 渤海;拟曳鳃虫属的南方拟曳鳃虫(见图)分布于东海陆架区。
渤海;拟曳鳃虫属的南方拟曳鳃虫(见图)分布于东海陆架区。
本词条内容贡献者为:
刘勇 - 副教授 - 西南大学资源环境学院
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国