

一、引言
内质网应激(ER stress)表现为内质网腔内错误折叠与未折叠蛋白聚集以及钙离子平衡紊乱 ,可激活未折叠蛋白反应、内质网超负荷反应和 caspase-12 介导的凋亡通路等信号途径 ,既能诱导糖调节蛋白 (glucose2regulated protein 78 kD , GRP78) 、GRP94 等内质网分子伴侣表达而产生保护效应 ,亦能独立地诱导细胞凋亡。 内质网应激直接影响应激细胞的转归 ,如适应、损伤或凋亡。
二、引起细胞ERstress的因素ER含有大量的伴侣蛋白、糖基化酶以及氧化还原酶为新生肤链的折叠提供了优化的环境,同时ER质量控制系统(ERqualityeontrolsystem)能通. 过ER相关降解作用(ER assoeiated degradation ERAD)降解非正确折叠的中间产物,凡影晌ER功能的因素都能够引起ERstress,包括有下列各种因素:
1.细胞营养物质缺乏包括葡萄糖饥饿和氨基酸饥饿,蛋白质及核昔酸的生物合成均需要必要的营养物质,所以葡萄糖饥饿和氨基酸饥饿代表一种代谢压力。
2.影响蛋白质翻译后修饰的因素,如还原物质二琉基苏糖醇(DTT)、β琉基乙醇(β-ME)、同型半胧氨酸(homoeystine);糖基化抑制剂衣霉素(tunieamyein)、葡萄糖胺(glueosamine)、2-脱氧葡萄糖(2-deoxyglueose)等。
3.影响ER钙离子平衡的药物,如ER Ca2+酶抑制剂Thapsigagrin,钙离子载体A23187,钙离子鳌合剂EGTA,抗生素lonomycin等。
4.突变基因表达的结构异常蛋白在ER堆积。
5.其它一些有害因素如细胞病毒感染等。
三、内质网应激激活的信号通路1.Irel通路哺乳动物细胞有2个Irelp同源物,Irelα和Irelβ,具有和Irelp相同的3个功能域, Irelα在各种细胞普遍存在,而Irelβ主要存在于消化道上皮细胞七。类似于酵母的Irelp, Irelα活化后,N端和BIP分离,c端具有核酸内切酶活性,能特异性地剪接转录因子xBPI的mRNA去除26个bp组成的片段,XBPI和HaCI的mRNA不具有同源性,但剪接机制类似。剪接后的XBPI mRNA具有活性,翻译产物与剪接前相比C端由于读码框移而改变。类似酵母,哺乳动物细胞的UPR靶基因含有保守的顺式作用元件ERSE,是由19个bp组成:ccAAT-Ng-ccAcG,当其ccAcG结合活化的xBPI或ATF6,同时ccAAT结合非特异性转录因子NF-Y,才能上调靶基因表达。有研究表明除了ERSE外,还存在功能类似的ERSEll(ATrGG-N-eCACG)。xBPI基因本身也含有ERSE,故剪接后的XBP-1蛋白除了能促进VPR靶基因的表达外,还能促进自身的表达。
-
PERK(PEK)通路PERK属于eIF2α蛋白激酶家族成员,和lrelα类似,是位于ER的I型膜蛋白,N端感受ERstress信号,存在非配体依赖性的二聚化结构域,非ERstress时二聚化位点被BIP遮盖,C端有丝/苏氨酸蛋白激酶功能域,但无核酸内切酶活性,PERK活化后能够特异性地磷酸化eIFZα的51位的丝氨酸,下调胞内蛋白合成的整体水平。当病毒感染或ER内钙离子耗竭引起ERStress时,存在于胞浆中的与PERK同源的蛋白激酶PKR也参与eIF2α的磷酸化。有研究表明,PERK活化后能特异性地抑制细胞周期素Dl的翻译表达,导致G.期的停顿。PERK磷酸化el兄。后抑制胞内大部分蛋白的翻译生成,同时也诱导三分之一的UPR基因的转录,其中包括XBPI基因,但机制并不清楚。
-
ATF6通路ATF6是位于ER的n型膜蛋白,哺乳动物细胞具有两种ATF6亚型ATF6α(90ku)和ATF6β(110ku),二者结构相似。N端是含有bZIP的转录激活功能域,C端位于ER腔内,具有多个BIP结合位点和两个高尔基体定位信号。非ERsrress时,ATF6和BIP形成稳定的复合物,通过BIP对GLS的抑制作用而停留在ER上。ATF6α和ATF6β在UΒR过程中通过相同的机制活化,ER腔内未折叠蛋白堆积信息能够使BIP和ATF6分离,BIP对GLs抑制作用的解除导致ATF6转移到高尔基体,由高尔基体蛋白酶S1P及S2P对其跨膜片段进行切割,产生游离的50kuN端片段,该过程类似于固醇反应元件结合蛋白(SRE-BΒ)的切割活化。活化的ATF6N端切割段可转移到核内促进含ERSE的转录因子(如XBPI)及UPR靶分子(如BIP)等基因转录,ATF6α和XBP1在功能上有所重叠,而ATF6日在UPR过程中并不起重要作用。有研究发现,除了ERsE外还存在有特异的ATF6结合位点(ATF6site):(G)(G)TGΑCGTG(G/Α),但ATF6Site在UPR过程中的作用并不很清楚,有趣的是ATF6site介导的基因转录可能完全依赖于Irel剪接系统,XBPI比ATF6有更高的ATF6Site亲和力,ATF6在NF-Y存在时对ERSE有更高的亲和力。
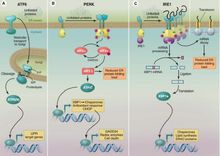
非折叠蛋白反应主要的三条通路
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国