

-壳1个,呈螺旋状,壳口大多具厣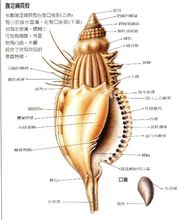
-头部明显,有眼及触角,口中有齿舌
-内脏团随螺壳的扭转一般呈螺旋形,左右不对称
-有的种类为卵胎生
-海产种类具担轮幼虫期和面盘幼虫期
起源与进化腹足纲动物包括我们熟悉的前鳃类的圆田螺(Cipangopaludina)、鲍(Haliotis);
后鳃类的壳蛞蝓(Philine)、海牛(Doris);肺螺类的蜗牛(Fruticicola)、蛞蝓(Limax)等。
这些动物的身体结构与原软体动物相比有很大不同。因为绝大多数的腹足类动物体外有一个螺旋卷曲的外壳,内脏器官失去了对称性,虽然它们的头与足还保持着对称。
人们从软体动物比较解剖学的研究知道除了腹足类动物外,其他各纲动物体制都是对称的,古动物学家从化石的研究也发现,从寒武纪早期的地层中的某些腹足类动物其贝壳也是对称的,例如一种化石腹足类Strepsodiscus壳对称呈平面盘旋。
以后人们在研究了腹足类的胚胎发育后也发现,腹足类的担轮幼虫也是对称的,而到了面盘幼虫后,身体突然出现扭转,随后是一个不对称的生长过程,最后成体变成了不对称的体制。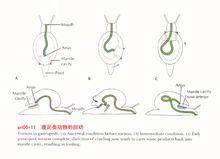
因此,从比较形态学、古动物学及发生学的研究都证明了腹足类动物早期的体制还是两侧对称的,而以后大多数种类的不对称是在进化过程中形成的。
那么在进化过程中腹足类是怎样形成了不对称的体制呢?人们推测腹足类动物也是由原软体动物一样的祖先进化而来,首先这种祖先动物由于体积的增加及头、足经常缩入壳下,使内脏囊得到了充分的发展并不断地在身体背部隆起,结果由内脏囊顶端悬垂下来的外套膜及由外套膜分泌的贝壳都随内脏囊的隆起而增加高度,使身体背部及贝壳成为长圆锥形,这种体形不利于动物在水中的运动及生存,于是逐渐地出现了由内脏囊的顶端开始做平面盘旋(coiling),后形成的外壳包围了先形成的外壳,这时壳与内脏囊仍是对称的,正如在早期地层中发现的化石种类壳是平面盘旋的那样。但这种平面盘旋使壳的直径很大,壳内的空间却很小,壳也不牢固,所以后期出现的壳不再是平面盘旋,而是沿一中心轴由上向下螺旋盘旋。
这样的螺旋壳的直径减少,壳内容积不变,但壳的牢度增加,壳又成为矮圆锥形,同时壳轴不再是垂直于身体的长轴,而是倾斜于身体长轴,使增大的内脏囊的重心移到了近前端以有利于运动。壳的螺旋盘旋或许说明了腹足类所以只有一对鳃、一对肾及一对收缩肌的原因。
壳的螺旋与倾斜虽然解决了内脏囊隆起所带来的问题,但结果外套腔被压在了壳下,水流的通畅、鳃的呼吸作用及排泄物、生殖细胞等的排出都受到了障碍,于是腹足类在进化中又出现了扭转现象(torsion),也就是说使身体扭转180°。当然这种扭转仅发生在外套膜及内脏囊部分,头、足不受影响,这种扭转或是顺时针方向或逆时针方向,扭转的结果使位于身体后端的外套腔移到了身体的前端;原来平行连接的侧脏神经索也因扭转而形成了8字形;左右侧的器官也交换了位置。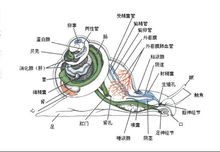
外套腔移到前端之后,水流、鳃、肛门、排泄孔及生殖孔都通畅了。但螺旋及扭转的结果,在以后的进化中,还是使一侧的器官发育受到阻碍,例如如果顺时针方向扭转,其壳口位于右侧,也称右旋,则其左侧的器官,鳃、心耳、肾得到发展,右侧的鳃、心耳及肾长期受阻以致最后退化消失,使身体只留下了一个鳃、一个心耳及一个肾,内脏囊由对称变成了不对称。如果逆时针方向扭转,其左侧的鳃、心耳及肾消失。
总之腹足类的体制变成了不对称,螺旋与扭转是两个过程,许多化石证据说明螺旋是先于扭转。在腹足类的进化中扭转过程是从寒武纪到奥陶纪内完成的,这一进化过程在海产腹足类的胚胎发育过程中得到了重演。还有一些腹足类动物在进化中经过了扭转之后,又发生了反扭转(detorsion),这就是后鳃类。
反扭转的结果外套腔又回到了身体的后方,而且大部分种类外套腔消失,鳃也消失,而出现了次生性的皮肤鳃,贝壳也逐渐退化。侧脏神经索不再成8字形,但由于扭转而消失的一侧器官(鳃、心耳、肾)不再因反扭转而恢复。随着不同程度的反扭转,外套腔与壳也出现不同程度的退化,到裸鳃目时,外套腔及壳、本鳃都已消失,身体又变成了蠕虫状,外表又出现了两侧对称。
至于肺螺类它们在进化中经过了扭转而没经过反扭转。由于陆地生活,它们的本鳃消失了,而由外套腔充血变成了“肺”进行呼吸,只有一个心耳及一个肾。由于侧脏神经节都移到前端食道周围,所以虽经扭转,侧脏神经索也不成8字形。扭转较之螺旋对腹足类动物有着更深刻的影响。
关于扭转的生物学意义有各种不同的看法,1972年Stasek提出扭转使外套腔移到身体前端,对头、足的缩入提供了空间,对动物起到了很好的保护作用。其次,鳃、嗅检器也随外套腔移到前端,这样不仅有利于气体的交换也使前端首先接触水流、感受水的性质。但肾及肛门移到前端易于造成对自身的污染,所以在平面盘旋的种,外套膜及壳的前缘背中线发生了裂缝或缺刻,以利于废物的排出及减少污染。
在螺旋卷曲的种类,外套缘在前端形成了出、入水管,以便于分清清洁的与污秽的水流,并可将废物更远的送出体外。
形态构造1、贝壳
壳顶、螺旋部、体螺层
螺旋纹、缝合线、螺轴、螺肋 前沟、后沟; 外唇、内唇 壳脐、壳口 方位
壳顶向上,基部向下,壳口向着自己,区分左右侧。
2、外套膜
3、头与足
头部:口(颚片与齿舌)、吻、眼、触角 足部
玉螺:前后足 蛇螺:无足 泥螺:侧足 鲍鱼:上下足 外套腔
4、厣:保护,足后端皮肤分泌,角质或石灰质。
5、内脏囊
6、消化系统 齿舌: 口 齿舌 消化腺 食道 肛门 胃
肉食性,少而有力,如脉红螺: 草食性:多而小,齿端钝,如鲍鱼
腹足纲动物身体也分为头、足、内脏囊及外套膜4部分。头部发达,具1~2对触手,触手的顶端或基部有眼。绝大多数种类体外有一发达的贝壳,头、足、内脏囊均可缩入壳内。腹足类的壳为典型的螺旋圆锥形壳,壳尖细的一端称壳顶(apex),是壳最先形成的部分,由壳顶围绕中心壳轴(columella)连续放大形成直径逐渐增大的螺形环,称为螺层(whorls),最后形成的一个螺层体积最大,称为体螺层(bodywhorl),其向外的开口即为壳口(aperture),头、足可由壳口缩入壳中。除体螺层之外,其他螺层称螺旋部(spire)。许多海产螺由壳口外翻,形成外唇(out lip)与内唇(inner lip)。壳表面有许多与壳口平行的细线称生长线(grow lines)。各螺层之间的交界线称缝合线(suture)。也有的种类壳口的前缘具水管凹陷(siphonal notch),或壳轴基部内陷形成脐(umbilicus)。如果以壳顶向前,壳口面向观察者,壳口在壳轴的右侧,则称为右旋壳(dextral shell),壳口在壳轴的左侧,称左旋壳(sinistral shell)。大多数腹足类动物为右旋壳。少数种为左旋壳,也还有少数种同时具有右旋壳个体与左旋壳的个体。
腹足类的壳因种不同,在形状、颜色、花纹及壳面装饰上表现出多样性。例如有的壳螺旋部不显著,成年的壳仅有体螺层极度膨大,如鲍(Haliotis),或壳形又表现出两侧对称,如帽贝(Limpet),或壳面长出骨刺,如骨螺(Murex),或壳完全埋在外套膜中,如壳蛞蝓(Philine),或壳完全消失,如海牛(Doris)等。壳的多样性不胜枚举。腹足类具有扁平、宽阔、适于爬行的足,类似于原软体动物。大多数前鳃类足的后端背面,有一圆形角质板,或石灰质板,它的大小、形状与壳口完全一致,当头、足缩回壳内之后,这个板十分严密的完全封闭壳口,此壳板称为厣板(operculum),具有保护作用。例如圆田螺的厣板是角质的,玉螺(Natica)、蝾螺(Turbo)是石灰质的,这是由于角质板上大量沉积的碳酸钙所致。
生活习性吸。
分布腹足纲种类多,世界上各个地带都有它们的踪迹。如有的生活在深海、浅海或潮间带,有的生活在江河湖沼,也有的生活在草原、森林、沙漠或高山。多数种类在陆地的浅水底下匍匐爬行,营自由生活;有些种类固着在岩石、珊瑚礁或其他贝壳上;有些种类在水中漂浮;也有些种类寄生在其他动物的体内或体外,营寄生生活或两者共生。
腹足类的运动腹足类的足富有肌肉质,适合于在各种条件下运动,但常与生活环境及生活方式相关。大多数地面或水底爬行的种类,是以足部肌肉的收缩来推动身体前进
。例如大蜗牛(Helix)运动时是先由足部的纵肌收缩,收缩到最高峰时,足底局部抬起离开地面,随后纵肌松弛,抬起的部分落回地面,这样便前进一步,如此的收缩波可以在足的局部范围内进行,也可在足的全长内发生,也可以左右交替收缩前进。纵肌的收缩由后向前推动身体前进,所以伸缩波与运动波方向一致。也有的种类靠足部伸肌伸长,然后横肌收缩从而拖曳身体前进,这样的伸缩波与运动方向相反。
一些小型生活在软质沙底的种类,可以靠足部纤毛运动推动身体前进,例如蜗牛、椎实螺(Lymnaea)等,其足部有丰富的腺体或腺细胞,它的分泌物在地面或植物上形成一层薄膜,再靠纤毛的摆动在薄膜上滑行,像扁形动物一样。还有一些在沙中穴居的种类,运动时靠足部充血形成犁或锚,然后拖动身体前进,如笋螺(Terebra)。还有一些水生后鳃类靠身体侧缘的波状收缩,或足部特化成翼在水中游泳运动。一般壳的螺旋部低平,在岩石上附着生活的种类不善于运动;
具有高螺旋部平行或倾斜于背部的壳,多为沙底生活的种类,一般较善于运动。许多海产种类,壳面上还有棘、刺等装饰物,以利于固着身体在泥沙中的位置。原始的种类还有一对壳轴肌(columella muscle),由足部伸到壳轴上,控制着足的伸出与缩回,但绝大多数的腹足类也由于身体扭转只保留一侧的壳轴肌。
水流与气体交换概述腹足纲动物中,绝大多数种类是水中生活,很少数为陆地生活。
水和气体可以不断的进入体内,腹足类就靠这种水流(也包括陆生种类的气流)完成身体的某些代谢作用。因为它们携带进新鲜的氧气,带走二氧化碳及含氮废物。各种腹足类动物由于外套膜、外套腔及鳃的结构不同,水流(或气流)经过身体的途径以及气体交换的方式也各不相同,这区种别在一定程度上反映了腹足类进化水平。腹足纲的前鳃类是比较原始的,是最先出现身体扭转的一类。
其中原始的种类,例如原始腹足目(Archaeogastropoda)还具有一对鳃,鳃均为双栉状(即鳃轴的两侧均有鳃丝),由于身体的扭转,外套腔移到了身体的前端,水由前端流入经过外套腔时,鳃在此进行气体交换,并将代谢产物等排出物随水流带出体外,为了避免排出物对鳃及外套腔的污染,一些种类在外套膜及壳的前缘中背处出现裂缝。
例如小裂螺(Scissurella),其肛门及肾孔开口在裂缝处,以避免污染。
又如潮间带生活的鲍,它是以外套膜及宽阔的形壳上形成一列小孔,以代替小裂螺的裂缝,其外套腔位于身体的左侧,鳃位于小孔之下,肛门及肾孔紧贴小孔,水流经过鳃之后再将排出物带走。
眼孔(Diodora)具有一个次生性对称的圆锥形壳,壳顶上有一小孔,外套膜由小孔处突出形成一水管,做为水流的出口。蝾螺及蜒螺(Nerita)具螺旋形外壳,但只保留了左侧的双栉鳃,水流由左侧流入外套腔,经鳃后再由右侧流出,肛门及肾孔开口在右侧外套膜边缘,排出物直接由出水流带走。这种由一侧流入、一侧流出倾斜于身体的水流方式能很好地解决了代谢产物的污染问题,因此大多数的前鳃类动物具有这种水流方式。
以上列举的这几种动物的不同水流方式,都属原始腹足目,它们主要限制在岩石表面生活,这与其鳃的结构相关。它们的鳃与原软体动物相似,鳃的背、腹面均有悬浮的膜以固定鳃在外套腔中的位置,鳃也都有较大的表面积,在水流通过时易于造成水流中颗粒物质的沉积而有碍鳃的呼吸,因此原始腹足目仅能生活在水流清洁、通畅的潮间带岩石的表面,而沉积物较多的软泥海底是找不到它们的分布的。
在前鳃亚纲中,原始腹足目仅是少数种类,绝大多数的种还是属于中腹足目(Mesogastropoda),例如圆田螺、及新腹足目(Neogastropoda)。
例如骨螺,在这两目中原始的双栉鳃已大部分消失(中腹足目中还有少数种保留),而代之以单栉鳃(unipectinate),仅鳃轴的一侧具有鳃丝;
另外外套膜前端延伸并卷拢形成水管(siphon),特别是在新腹足目,更为普遍,它们可以通过出水管将代谢物带走,水流在体内仍保持着与蝾螺等相似的倾斜水流,这种鳃的结构、水管及水流方式适合于沙质水底表面生活或穴居,大量的海产前鳃类都是这种生活方式。
极少数的种类侵入淡水甚至陆地,总之凡是淡水生的皆为单栉鳃,倾斜水流,如果侵入陆地则鳃消失而由外套膜进行皮肤呼吸,并有厣板减少水分的丧失。
后鳃类来自于前鳃类,在进化过程中,由于它们又经过了反扭转,其外套腔又移到了身体的右侧或靠近身体的后端,这样由于外套腔移到身体前端所带来的污染问题也就不存在了。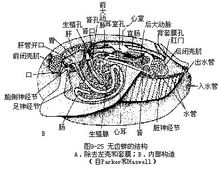
虽然反扭转的原因尚不清楚,但由反扭转而出现的一种趋势是明显的,即壳逐渐减少及丢失,外套腔及鳃也退化及消失,身体由不对称又重新出现了两侧对称。在最原始的后鳃类,例如捻螺(Acteon)仍具有圆锥形壳,仍有一单栉鳃,它的水流及呼吸方式仍相似于前鳃类。在较高等的海兔(Aplysia),其壳减小被包在外套膜中,外套腔及鳃仍然存在,但大大减小。而裸鳃目动物中,例如蓑海牛(Aeolis)的壳、外套腔、鳃均已消失,而代之以身体的皮肤形成许多突起形成一种次生性的皮肤鳃也称为裸鳃(cerata),以进行呼吸。这种皮肤鳃有的像蓑海牛一样是分布在身体整个背面,有的分布身体的某一定部位,如舌尾海牛(Glossodoris)的这种鳃是在身体背面后端围绕肛门分布,总之裸鳃目动物的本鳃消失,代以皮肤鳃进行呼吸。

许多种胚胎期中有厣板而成年期无厣板,由此可知此类是来自前鳃亚纲,并包括相当多成功的陆生及淡水生活的种类。
它们大多数为右旋壳,除了身体右侧有外套膜形成的一个气孔(pneumostome)之外,外套腔已完全封闭,本鳃消失,而是由外套膜壁高度充血形成血管网,所谓“肺”,其实是靠外套膜底部的搧动以造成气体在外套膜腔中的流通,肺螺类即用这种方式进行气体交换,吸收空气中的氧,即使水生种类也是如此。
肺螺类中仅有很少的种是海产的,且多生活在潮间带或河口处,多数种类分布在亚热带及温带地区的陆地及淡水中。低等的肺螺类,如基眼目的椎实螺,为水生的种类,但它们不能吸收溶于水中的氧,而必须周期性的到水面上进行气体交换,用外套膜形成的水管伸到水面,交换气体后关闭气孔再沉入到水下,在水下停留几分钟到十几分钟因种而不同,然后再浮到水面上。还有的水生种类可以在气孔处由外套膜褶雏形成次生性的鳃进行呼吸,例如扁卷螺(Planorbids)就是这样。
在较高等的柄眼目,例如大蜗牛(Helix)及蛞蝓(Limax)等,它们是陆生种类,壳很薄,其外表有较完整的角质层,具有很好的拒水性,壳均为小型,直径小于1cm,但也有些种的壳完全包埋在外套膜中或完全消失,而仍有气孔,例如蛤蝓的壳已退化成一极薄的石灰质板。甚至有些种外套腔、贝壳完全退化消失,例如石蟥(Oncidium)(石蟥为海滨生活)。总之陆生的肺螺类多生活在石块、树皮下,或潮湿的地面,气孔也经常关闭,以避免由于呼吸作用而使体内水分散失。当气候干燥或炎热的季节,它们甚至躲在阴湿的环境下分泌粘液封闭壳口,以度过恶劣环境。
除了肺螺之外,前鳃亚纲的个别种,也能形成类似的“肺”进行气体交换,例如钉螺(Oncomelania),这是一种水陆两栖的螺,陆生时即用肺呼吸
腹足类动物所以种类繁多,分布广泛,是与其具有多种食性及取食方式相关的。
腹足类动物包括有植食、肉食、腐食、悬浮取食,以及寄生等多种不同的食性。
植食性包括前鳃类中许多海产及淡水生活的种类、绝大多数的后鳃类及肺螺类。植食性是腹足类中最重要的取食习性。
植食性的腹足类是以各种海藻,水生或陆生植物为食,根据不同的生活环境而取食不同的植物,有些种甚至造成农业上的危害。
腹足类是以齿舌做为刮取食物的器官,齿舌的取食方式也因种不同,具有锉、刷、刀及运输器官等多种机能。齿舌上有成排的细齿,每个横排细齿的数目可由几个到几百个,分为中央齿(central tooth),通常一枚、其两侧为侧齿(lateral teeth)一对到许多对,再两侧为一对到许多对边缘齿(marginal teeth)。每种细齿的形状、数目及排列等特征为分种属的依据之一。
在植食性种类中,原始的种类齿的数目多,结构简单一致呈扇形排列,肺螺类的大蛞蝓的齿数可多达26 800个。高等的种类齿的数目减少,齿的顶端还有锯齿。
齿舌回收时,边缘齿可以收集食物,再经齿舌带送入食道。在一些后鳃类及肺螺类口腔内除了齿舌之外,还有颚(jaw),它是一种角质结构,以帮助齿舌切割食物。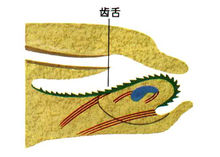
它们的食道或胃的前端变成了嗉囊(crop)及砂囊(gizzard),例如海兔嗉囊是食物暂时贮存的部位,而砂囊内常裹有几丁质(海兔)或含有许多砂粒(淡水螺)以帮助研磨食物。陆生的种类没有砂囊。其消化作用由嗉囊进行,在一定程度上代替了胃的作用,例如蜗牛。
食道之后即为胃,是食物消化的主要场所。
在前鳃亚纲中,由于扭转,胃的位置也倒转,即食道在胃的后端进入,肠由胃的前端通出。在原始的种类,例如原始腹足目的许多种,胃与原软体动物相似,胃内仍保持有晶杆囊、晶柱、胃楯、筛选区等结构,消化道也较长,仍行胞外消化与胞内消化。在较高等的种类,如中腹足目及新腹足目,胃已失去了原始结构,而或多或少的变成了一个简单的囊,已完全行胞外消化,如田螺,并具有发达的肝脏。
胃接受肝脏及各种消化腺所分泌的酶进行食物的消化,其消化酶主要是淀粉酶及纤维素酶。胃后即为肠,在植食性的种类,肠道一般很长,而且盘曲,肠壁向内突起形成嵴与沟。腹足类其食物在消化道中的运行,特别是在胃与肠中的运行是由纤毛作用的推动而不是靠肌肉的收缩。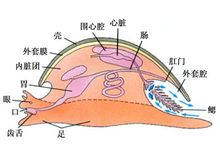
在原始的种类,胃壁内有筛选区等结构,肠壁内也有一系列的嵴与沟,但这些嵴与沟结构简单。在较高等的种类,胃壁及肠壁内的嵴与沟也较复杂。沟与嵴的表面布满了纤毛。由唾液腺分泌物粘成的食物索进入胃内后,由消化酶将粘液除去,又形成食物颗粒,这些食物颗粒在经过胃、肠的嵴与沟时,按颗粒的大小进行了分类,微小的颗粒落入沟内,再由沟内的纤毛摆动将颗粒向后运行,胃壁筛选出的微小颗粒还可进入消化盲囊中进行胞内消化及吸收。消化道内较大的颗粒只能在嵴上向后运行或在胃内进行胞外消化或进入肠道,不能消化的食物颗粒形成粪便经肛门开口在外套腔排出体外。在进化的种类由于沟与嵴的复杂化,可以同时筛选分离出4种或更多种大小不同的颗粒,分别在不同的沟与嵴中运行。
食物在消化道中的这种运行方式可见于所有的软体动物中。
肉食性腹足类后鳃亚纲及肺螺亚纲极少数为肉食性的。其消化道的前端常形成吻,周围包有吻鞘,吻中含有口腔、齿舌。取食时是由于头部充血而将吻伸出,缩回时是受肌肉控制,由缩吻肌将吻鞘拉回。
肉食性种类的齿舌上细齿的数目减少,但硬度增加,齿舌末端有倒勾、棘等,适合于切割及撕裂食物,有时齿上还有毒腺。一种肉食性的芋螺(Conus),齿变成了单针状,中央具沟,末端具倒刺、基部有毒腺,其分泌物可以麻醉及杀死捕获物。肉食性种类口腔中常有颚,消化道一般较植食性的短,消化腺分泌的酶主要是蛋白酶。还有一些肉食性腹足类具有钻孔取食的习性。
例如中腹足目的玉螺(Natica)、乳玉螺(Polinices),新腹足类的骨螺、红螺(Rapana)等,其足的前端有一个可外翻的腺体,它可以分泌酸性物质,以溶解壳的有机质及钙质成分。这些动物取食其他软体动物时,先用齿舌研磨捕获物的外壳,再由腺体分泌酸性物,如此反复多次,直到壳被穿透之后再用吻取食或用颚取食。
在肉食的裸鳃目中一些种具有很有趣的生物学现象,它们在取食腔肠动物之后,能将腔肠动物的刺细胞通过胃、盲囊最后贮存在背部的皮肤中,用来做自己的防卫工具。
当这些腹足类遇到敌人时,可以将刺细胞放出,其排放的机制是肌肉收缩产生的压力而放出,刺细胞被排放之后可再从食物中得到补充。不同种的裸鳃类,有固定的刺细胞,可能是由于裸鳃类取食不同的腔肠动物所致。
腐食性或沉积取食者例如织纹螺(Nassarius)、觿螺(Hydrobia)等,它们大量的出现在潮间带,数量多时每平方米达到三万头觿螺。
它们以潮汐带来的沉积物碎屑或腐烂的有机物颗粒为食。一些生活在淤泥较多的地方也多有这种取食方式。
还有一些腹足类为悬浮取食者,例如履螺(Crepidula)它们的鳃丝高度延伸以增加鳃的表面积,以便于捕捉各种浮游生物或微个的食物颗粒,其外套腔很小,仅在身体前端留有小缝,水由左侧流入再由右侧流出,随水流带入外套腔的浮游生物被鳃丝上的粘液捕获,然后送入口内。一些管居的蠕虫状的螺类,例如小蛇螺(Serpulorbis)可以利用足腺分泌粘液,粘液排出壳外被波浪作用分散成许多细粘液丝,分布在壳口周围,藉以诱捕浮游生物。相邻的个体之间所分泌的粘液丝互相连接,形成诱捕网联合诱捕。
有趣的是悬浮取食的与沉积取食的后鳃类胃又重新出现了晶杆囊与晶杆等结构,它们的形态与双壳纲的晶杆十分相似,这可能是由于它们的取食习性及取食方式相似所造成的次生现象。腹足纲中还有极少数种类为寄生生活,并经过外寄生发展到某些种的内寄生。
外寄生外寄生的种类仅涉及口区与消化道的改变,例如短口螺(Brachystomia),小塔螺(Pyramidella),它们具有几丁质的颚,具有泵状咽及晶杆胃,以适合于吸食多毛类及双壳类动物的体液,寄生螺(Stylifer)是寄生于棘皮动物体壁上的一种小型螺,其壳仍存在,足已高度退化,它的吻特别发达,以利于吸食寄主的体液及组织。改变最大的还是内壳螺(Entoconcha),它寄生于海参体内,身体已改变成蠕虫状,贝壳仅在幼虫期出现,以寄主的体液为食。Enderoxenos的消化道已完全退化消失,靠体表吸收寄主的组织为食。
腹足纲动物的排泄器官是后肾,肾的数目与鳃及心耳是一致的,只有在原始腹足目还保留有一对肾(也有一对鳃),并且具有一对体腔管(围心腔管)(renopericardial canal),做为排泄管,其一端与围心腔相通,一端开口到外套腔,生殖腺也开口在排泄管中,所以它兼有生殖导管的作用。
随着原始腹足目水流方向与鳃部位的改变,它们的后肾,体腔管及生殖腺发生着改变,即后肾由二个变为一个,生殖腺通过体腔管到独立开口于外套腔。在中腹足目及新腹足目及后鳃类等,由于身体扭转,使一侧的肾消失,只保留了一个肾,且肾囊的内壁向内发生许多褶皱以增大排泄的表面积,肾管直接开口外套腔,体腔管消失。生殖腺也独立开口在外套腔中。
腹足类的排泄是由后肾及体腔的过滤作用及后肾的重吸作用而完成。后肾除了从血液中收集代谢产物也由围心脏收集废物,由于各种腹足类生活环境的不同,其排出的废物也不同。水生种类特别是淡水种类,其排出物主要是氨(ammonia),氨对组织有较高的毒性、溶于水,因此在氨形成及排出时都需要伴随一定的水分;后肾对排出物中的盐及有用物质回收后,将它们变成低渗的含氨的尿经肾孔开口到外套腔排出体外。
陆生种类其排泄物是尿酸(uric acid),尿酸不溶于水,它对组织的毒性较低,且不需要伴随水分排出,因此可减少体内水分的丢失;特别在一些陆生种类其运动方式为纤毛滑行(如蜗牛),其足必须先分泌大量的粘液;这样便会丧失大量的水分,所以由陆生种类的排泄物中减少水分的丢失这是十分重要的,是它们对陆地生活的一种适应。
另外陆生螺类生活在潮湿环境,昼伏夜出,当恶劣环境时便进入夏眠(estivation)或冬眠(hibernation)也都是为了减少体内水分的丧失。此外大多数腹足类的消化腺也有一定的排泄作用,因为消化腺中的某些细胞具收集代谢产物的功能,其收集物最后通过肠道排出体外。
腹足类的循环腹足类的循环系统为开放式。
前鳃亚纲由于体制扭转,围心腔及心脏位于内脏囊的前端,在原始腹足目中一些种类仍然具有二个心耳。它们的循环系统的结构及途径相似于原软体动物。
其他的腹足类动物心脏或在内脏囊的前端或在其后端,但只有一个心耳。在循环过程中形成发达的血窦。由心脏向前端分出动脉到头与足,向后分出动脉到内脏。
在前端血液由小血窦汇合成发达的头足窦(cephalopedal sinus),身体后端的血液汇合成肾窦(subrenal sinus),以后这两个血窦联合,经肾脏再进入鳃中进行气体交换,血液出鳃后再流回心耳及心室,心室的收缩是靠其中肌浆蛋白质的收缩。肺螺类由于鳃的消失,气体在肺的血管网处交换后直接流回心耳及心室。
前鳃类及肺螺类动物的血浆中,一般都含有呼吸色素即血蓝素(hemocyanin),这是一种含铜的蛋白质。淡水生活的扁卷螺类(Planorbidae)(肺螺类)含有血红素(hemoglobin),它是一种含铁的蛋白质,这些呼吸色素易于与氧结合及释放氧,有利于呼吸。
对后鳃类的呼吸色素知道的很少,但对广泛研究过的海兔知道它是没有呼吸色素的。
腹足类的神经系统包括脑、足、侧、脏4对基本的神经节。
脑神经节(cerebral ganglion)位于食道背面,由它发出神经支配触手、眼等;又分出一对口球神经节(buccalganglion),支配口腔及齿舌的运动。
足神经节(pedal ganglion)位于足中部肌肉中,支配足的运动,且有神经索与脑神经节相连。侧神经节(pleural ganglion)位于脑后食道两侧,主要支配外套膜及闭壳肌的收缩,也有神经索与脑相连。脏神经节(visceral ganglion)位于内脏囊内,侧脏神经节之间有神经索相连,侧脏神经索上还有周缘神经节(parietal ganglion),它支配着肠的运动。
但在前鳃类,侧脏神经索由于扭转而形成8字形。后鳃类由于反扭转,侧脏神经索又平行排列失去8字形。肺螺类的4对基本神经节均集中在头部,侧脏神经索消失,不表现出扭转的影响。感官包括触手、眼、嗅检器及平衡囊。前鳃类具有一对触手位于头的前端,大多数后鳃类及肺螺类具有两对触手。
触手上分布有化学感觉细胞及触觉感觉细胞。在一些原始的前鳃类成员,足及外套膜的边缘可以分布有许多小触手,其功能也是与头端的触手相似。眼是其重要的感官,位于触手的基部(一对触手)或位于后触手的顶端(二对触手)。原始的眼仅包括色素细胞及光感觉细胞,排成一杯状,在高等的种类眼呈球形,眼的表面还有角膜(cornea),内有晶体(lens),这种眼也只能感光而不能成像。
嗅检器与鳃密切相关,其数目与鳃的数目是一致的,位于外套膜表面靠近鳃的部位,鳃消失的种类,其嗅检器也消失。嗅检器是一种化学感受器,它对水中的沉积物及水质的变化很敏感,所以肉食及腐食性种类嗅检器较发达。腹足类具有一对平衡囊,位于足内靠近足神经节的地方,有感受平衡的作用,固着生活的种类,平衡囊不存在。
生殖与发育腹足类为雌雄异体或雌雄同体,其生殖腺(精巢、卵巢或精卵巢、也称两性腺)位于内脏囊靠顶端,在原始腹足目中许多种类的生殖细胞由肾管排到外套腔,再随水流到外界,受精作用在海水中进行。
其他腹足类的生殖导管独立于肾管,但从发生上也是来自于体腔管。前鳃类多为雌雄异体,雄性个体包括精巢与输精管,输精管的后端有前列腺以产生授精液,输精管的末端形成交配器官阴茎(penis),前列腺开口在阴茎内,其授精液帮助交配。
雌性个体有卵巢、输卵管,输卵管可膨大形成蛋白腺(albumin gland)、受精囊(seminal receptacle)及粘液腺(mucous gland)。蛋白腺及粘液腺可分泌营养物及粘液,以形成卵膜及卵囊,输卵管的末端还伸出一交配囊以贮存交配后的精子。雌雄性需交配后方能使卵受精。
少数种类为卵胎生(ovoviviparity),例如圆田螺,为雌雄异体,其雌性输卵管的末端膨大形成子宫,卵在此发育成小田螺后才经生殖孔排出体外。
后鳃类、肺螺类及极少数的前鳃类为雌雄同体。其两性腺或同时形成精子与卵子或雄性先熟先形成精子。生殖细胞成熟后经过一段两性管,然后雌、雄生殖管分开并行。两性管的末端有一受精囊,为卵受精的部位。受精囊的一侧有蛋白腺,它分泌蛋白质及粘液包围受精卵。雄性生殖管末端膨大形成阴茎,用以交配,雌性生殖管分出具长柄的交配囊,粘腺及射囊(dart sac),交配囊贮存交配后的精子,粘腺用以包围卵,射囊的分泌物形成射针,用作性刺激物,输卵管的末端形成阴道。它们也需要交配受精。
水生的种类交配时常只有一种生殖腺先成熟,它们或作为雌体,或作为雄体进行交配。陆生种类交配时,常是互赠精囊到对方,交配后,精囊到对方的交配囊中释放出游离的精子,然后精子游到受精囊中使卵受精。产卵时卵的形状、数目、卵囊形态等随种而异,卵多形成卵囊。水生种类产卵在水草或其他物体上,陆生种类产卵在潮湿土壤、洞穴等阴湿地方。
腹足类均为螺旋卵裂,经内陷法与外包法形成原肠胚,仅在原始腹足目中存在独立的担轮幼虫期,经一段时间的游泳后变为面盘幼虫。其他种类的腹足类没有独立生活的担轮幼虫期,在卵内已渡过担轮幼虫期。海产的种类绝大多数都有自由游泳的面盘幼虫期,它具有两个大的纤毛叶,是来自担轮幼虫期的口纤毛环,具有运动与取食的作用。
面盘幼虫期已出现了足、触手、眼及壳,在面盘幼虫后期出现了扭转,这一过程可能在数分钟内或数日内完成,因种而不同。足生长迅速,用以爬行,以后用足附着在底部变态成成体。后鳃类的发育中还经过了反扭转。肺螺类没有自由生活的幼虫期,孵化时已为幼年成体。1
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国