

遗传信息(genetic information) 指生物为复制与自己相同的东西、由亲代传递给子代、或各细胞每次分裂时由细胞传递给细胞的信息, 即碱基对的排列顺序,或指核苷酸的排列顺序,DNA中的脱氧核苷酸、RNA中的核糖核苷酸的排列顺序。1
研究领域遗传信息是指生物为复制与自己相同的东西、由亲代传递给子代、或各细胞每次分裂时由细胞传递给细胞的信息,即碱基对的排列顺序(或指DNA分子的脱氧核苷酸的排列顺序) 。
在1866年首先是由G.J.Mendel(1866)的研究形成了概念,即相应于生物各种性状的因素(称为基因)中包含着相应的信息(以后G.Beadle等人(1941)所开创了遗传生物化学的研究,描绘出这样一个轮廓:基因和决定生物结构与功能的蛋白质之间具有一对一的对应关系。1
关于基因的化学本质方面,根据O.T.Avery等(1944)进行的转化实验,以及A.Hershey和M.Chase(1952)用大肠杆菌噬菌体的DNA进行的性状表达实验,已阐明DNA是遗传信息的载体。附着DNA结构研究的进展,1960年已经确立了这样的概念,即基因所具有的信息可将DNA的碱基排列进行符号化。信息在表达时,DNA的碱基排列首先被转录成RNA的碱基排列,然后再根据这种排列合成蛋白质。有的病毒的遗传信息的载体不是DNA,而是RNA。遗传信息不仅有相应于蛋白质的基因信息,也包括对信息解读所必需的信息、控制信息表达所必需的信息,以及生物为了复制与自己相同结构所必需的一切信息。1
种类基因中脱氧核苷酸种类、数目和排列顺序的不同,决定了遗传信息的多样性。
遗传传递分类生物体遗传信息的传递的几种类型;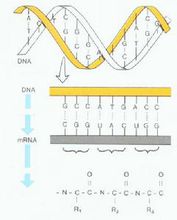
中心法则及其补充内容告诉了我们遗传信息的流动方向。其分解过程包含了如下6点:DNA的复制,遗传信息流动方向由DNA→DNA;DNA的转录,遗传信息流动方向由DNA→RNA;翻译,遗传信息流动方向由RNA→蛋白质;RNA的复制,遗传信息流动方向由RNA→RNA;RNA的逆转录,遗传信息流动方向由RNA→DNA;蛋白质的复制,遗传信息流动方向由蛋白质→蛋白质。但是究竟在生物体中遗传信息的传递应该包含其6点内容中的几种呢?不同类型的生物,遗传信息的传递过程也有所差异。生物体遗传信息的传递大致分为如下类型:
DNA复制型
在DNA复制型的生物中,生物体的遗传信息流动包含3点:DNA的自我复制,遗传信息流动方向由DNA→DNA;DNA的转录和翻译,遗传信息流动方向由DNA→RNA→蛋白质。这种类型的生物主要针对地球上绝大多数的动植物和噬菌体病毒等。1
RNA复制型
在RNA复制型的生物中,生物体的遗传信息流动包含2点:RNA的自我复制,遗传信息流动方向由RNA→RNA;翻译,遗传信息流动方向由RNA→蛋白质。这种类型的生物主要针对植物病毒如烟草花叶病毒和动物病毒如脊髓灰质炎病毒等。也有些遗传信息的流动只有1种:RNA的自我复制,遗传信息流动方向由RNA→RNA;这种类型的生物主要针对SARS病毒,流感病毒等。
RNA逆转录型
在RNA逆转录型的生物中,生物体的遗传信息流动包含3点:RNA的逆转录,遗传信息流动方向由RNA→DNA;转录,遗传信息流动方向由DNA→RNA,翻译,遗传信息流动方向由RNA→蛋白质。这种类型的生物主要针对致癌病毒和导致艾滋病的人体免疫缺陷病毒(HIV)。
蛋白质复制型
在蛋白质复制类型的生物中,生物体的遗传信息流动包含1点:蛋白质的复制,遗传信息流动方向由蛋白质→蛋白质;这种类型的生物2000年以前只发现一种即盛行欧美的疯牛病病毒(朊病毒)。
起源遗传信息是由三联体密码子记载的,因此遗传信息的起源归根结底就是密码子的起源问题。迄今为止,提出了若干假说。这些学说分别从偶然性、化学相互作用、协同演化、生化系统起源以及综合作用等不同视角探讨了遗传密码子起源的可能途径,特别是以生化系统构建为目的的从能量转化到信息化的演化机制值得关注。2
凝固事件假说克里克(Francis Crick)提出凝固事件假说(frozen accident hypothesis),认为密码子与氨基酸的关系是在某一时期固定的,之后很难再被改变。现在所有的生物几乎使用着同样一套密码似乎支持这一假说,这也表明,所有生物起源自单一的共同祖先(Crick 1968)。这推测了演化事件的重要时间节点。3
立体化学假说韦斯(Carl Richard Woese)提出了立体化学假说(stereochemical hypothesis),认为氨基酸与它们相对应的密码子有选择性的化学结合力,即遗传密码的起源和分配与RNA和氨基酸之间的直接化学作用密切相关,或者说,密码子的立体化学本质取决于氨基酸与相应的密码子之间物理和化学性质的互补性(Woese et al. 1966)。这可能是密码子起源的一个重要化学机制。4
Polyansky等(2013)通过实验和计算发现,mRNAs中不同核酸碱基的密度分布,非常类似于它们所编码的蛋白质中这些相同核酸碱基的氨基酸亲电子密度分布,遗传密码进行了高度最佳化,以最大化这种匹配。
共进化假说王子晖(J. Tze-Fei Wong)提出了共进化假说(co-evolution hypothesis),认为密码子系统有原始氨基酸形成的前生物代谢途径的印迹,因此可以从氨基酸代谢途径发现密码子的演变过程,即密码子的进化与氨基酸生物合成的进化是并列的。该假说认为,氨基酸和相应编码的忠实性反映了氨基酸生物合成路径的相似性,并非物理化学性质的相似性(Wong 1975)。这推测了密码子起源的一种可能路线,但是,从简单的原料合成各种氨基酸可能是发生在前生命演化末期的事情了。5
ATP中心假说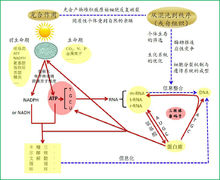 谢平(2016)提出了ATP中心假说(ATP-centric hypothesis),认为遗传密码子是生化系统的一部分,因此,必须与生化系统的演化相关联,而生化系统的核心是能量载体——ATP(各种生化循环,如卡尔文循环、糖酵解和三羧酸循环等,均与ATP相耦联),它是光能转化成化学能的唯一终产物,它可以衍生出其它核苷酸,这些核苷酸可以自身缩合成核酸。ATP可活化氨基酸,为多肽链的缩合提供了能量基础。因此,只有ATP才能建立起核酸和蛋白质之间的联系。6
谢平(2016)提出了ATP中心假说(ATP-centric hypothesis),认为遗传密码子是生化系统的一部分,因此,必须与生化系统的演化相关联,而生化系统的核心是能量载体——ATP(各种生化循环,如卡尔文循环、糖酵解和三羧酸循环等,均与ATP相耦联),它是光能转化成化学能的唯一终产物,它可以衍生出其它核苷酸,这些核苷酸可以自身缩合成核酸。ATP可活化氨基酸,为多肽链的缩合提供了能量基础。因此,只有ATP才能建立起核酸和蛋白质之间的联系。6
综合进化假说Knight等(1999)提出了综合性假说,认为遗传密码是由选择(selection)、历史(history)和化学(chemistry)三个因素在不同阶段起作用的。该假说认为,在遗传密码起源初期,可能主要是氨基酸和密码子之间的直接相互作用来决定氨基酸的编码;在新氨基酸的引入和密码子扩展阶段,共进化作用可能占据主导地位;而随着tRNA的进化和蛋白质的功能增加,逐渐去除了氨基酸和密码子的直接相互作用,密码子在不同尺度上的交换在某些程度上允许通过密码子的重新分配进行优化。7
其它假说1981年艾根提出了试管选择(in vitro selection)假说,1989年英国化学家奥格尔(Leslie Eleazer Orgel)提出了解码(decoding)机理起源假说,1988年比利时细胞生物学和生物化学家杜维(Christian de Duve,1974年获诺贝尔生理学或医学奖)提出了第二遗传密码(second genetic code)假说。
英国巴斯大学的Wu等(2005)推测,三联体密码从两种类型的双联体密码逐渐进化而来,这两种双联体密码是按照三联体密码中固定的碱基位置来划分的,包括前缀密码子(Prefix codons)和后缀密码子(Suffix codons)。不过,也有人推测三联体密码子是从更长的密码子(如四联体密码子quadruplet codons)演变而来,因为长的密码子具有更多的编码冗余从而能抵御更大的突变压力(Baranov et al. 2009)。
2007年中国科学院北京基因组研究所的肖景发和于军(2007年)提出了遗传密码的分步进化假说(stepwise evolution hypothesis),认为最初形成的遗传密码应该仅仅由腺嘌呤A和尿嘧啶U来编码,共编码7个多元化的氨基酸,随着生命复杂性的增加,鸟嘌呤G从主载操作信号的功能中释放出来,再伴随着C的引入,使遗传密码逐步扩展到12,15和20个氨基酸(肖景发和于军2009)。
厦门大学的有机化学家赵玉芬(1994-1996年)也曾提出核酸与蛋白共同起源的观点,认为“磷是生命化学过程的调控中心”,因为磷酰化氨基酸能同时生成核酸及蛋白,又能生成LB-膜及脂质体。她认为,原始地球火山频发,焦磷酸盐、焦磷酸脂类化合物容易在地表积累,其P—O—P键含有的能量,通过与氨基酸形成P—N键,最终转移到肽键和核苷酸的磷酸二酯键中。她推测,磷酰化氨基酸在同时生成蛋白质和DNA/RNA的过程中,蛋白质与DNA/RNA可以通过磷酰基的调控作用相互影响,从而产生了原始密码子的雏形,并进一步进化到遗传密码的现代形式。但问题是,磷酰化氨基酸为何要导演核酸和蛋白质的共进化故事呢?
也有将关于密码子起源的各种学说分为这样四类的:化学原理(Chemical principles)、生物合成扩展(Biosynthetic expansion)、自然选择(Natural selection)和信息通道(Information channels)。根据信息理论研究中的率失真模型(rate-distortion models)推测,遗传密码子的起源取决于三种相互冲突的进化力量的平衡:对多样的氨基酸的需求、抵御复制错误以及资源最小成本化(Freeland et al. 2003,Sella and Ardell 2006,Tlusty 2008,)。6
本词条内容贡献者为:
李少斌 - 副教授 - 长江大学生命科学学院
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国