

泰坦巨龙类(Titanosauria),也称为泰坦龙类、巨龙类,是蜥脚下目的一个演化支,包括萨尔塔龙与伊希斯龙,也包括一些地球上最重的陆行动物,像阿根廷龙、潮汐龙;的体重可达100公吨(虽然存在着许多质疑)。古生物学家以古希腊神话中的巨人族泰坦命名这个超科,泰坦神族的地位高于奥林匹斯十二主神。 泰坦巨龙类与腕龙科等相近物种,属于范围较大的巨龙形类演化支。
简介主要于白垩纪期间在南半球繁盛的蜥脚类,有像甲龙那样拥有铠甲等各种各样的种类,大小也各不相同。口部前端有长长的棒状牙齿,鼻孔开口于头顶附近。此外,脊椎骨的棘突较低,髂骨的前方分别向左右分开,所以背部和腰部非常宽广为其特征。泰坦巨龙类包括稍显原始的安第斯龙科和泰坦巨龙科。因为完整的骨路化石很少,所以还留有很多谜团1。
它们主要活跃在白垩纪的南半球,是一群憨厚可亲的恐龙族群,但若以为它们天性温善好,那就大错特错,只要内食龙胆敢把它们逼火,它们也会团结起来与之拼个鱼死网破,你死我活,泰坦巨龙类中既有像甲龙类带有青背甲的恐龙,也包含若体形大小不一的恐龙,形态多种多样,大都口中长有像谷穗一样的长牙,鼻孔位于脑门前面,它们常以骨突脊低、解骨前端左右开叉使得背部面和腰为宽广为其特征。一般包含着安第斯龙科和泰坦巨龙科中的恐龙,但在森坦巨龙类中,完的恐龙化石为少见,因而圣今仍然存在着许多未解之谜2。
特征泰坦巨龙类的头部比较宽,类似圆顶龙与腕龙,不过巨龙类的头部比较长,而与其他蜥脚类比起来,泰坦巨龙类的头部显得比较小。它们属于大鼻龙类,拥有大的鼻孔,而且鼻骨是隆起的,牙齿小型且成铅笔状或钉状。以蜥脚类而言,泰坦巨龙类的颈部是比较短的。尾巴类似一条鞭子,但没有梁龙科的尾巴长。泰坦巨龙类的骨盆是比其他蜥脚类纤细,而胸部是比较宽的,让泰坦巨龙类的四足呈现独特的步态,因此它们的脚印是比其他蜥脚类来的宽广。泰坦巨龙类的前肢是比后肢来的粗短,脊椎是实心而非空心的,这也许是一个倒退回原始蜥脚类的特征。不过它们的脊椎组织是更有弹性,所以泰坦巨龙类可以比其他的蜥脚类移动的更迅速。
从化石的皮肤痕迹发现,在它们皮肤上大的鳞片周围镶嵌著较小的鳞片。古生物学家甚至在萨尔塔龙的皮肤上发现类似甲龙的坚甲。
虽然泰坦巨龙类恐龙的体型巨大,但有些物种体型较小,可能是因为异域种化与岛屿环境隔离而造成的侏儒物种,例如马扎尔龙。

食性在晚白垩纪时期的泰坦巨龙类粪化石中发现了植物岩(Phytoliths)与硅化的植物碎片,显示了泰坦巨龙类广泛且不挑剔的食性。一个在2005年公布的发现,显示泰坦巨龙类有广泛的进食范围,除了预期中的苏铁与针叶树,还有单子叶植物,包括棕榈科、禾本科、稻米与竹的祖先;有假设认为这些新植物造成草食性恐龙与草的协同进化。
蛋巢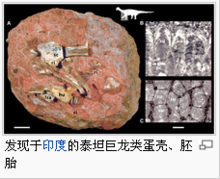
最近在阿根廷巴塔哥尼亚的Auca Mahuevot发现了一个大型的泰坦巨龙类蛋巢,在西班牙也有类似的发现。这些小型恐龙蛋的直径为11到12厘米,其中包含化石化胚胎,以及完整的皮肤压痕,但无法显示是否有羽毛或皮棘。很明显地有数百只萨尔塔龙科雌性个体挖掘洞穴,将蛋产于洞中,并用泥土与植被覆盖恐龙蛋。这些证据显示泰坦巨龙类是群体动物,身体拥有装甲,这些装甲以及群体行动可能用来抵抗大型掠食动物的攻击,例如:阿贝力龙。
地理与年代范围泰坦巨龙类是白垩纪-第三纪灭绝事件之前最晚出现的大群蜥脚下目恐龙,生存于9,000万年前到6,500万年前,是当时的优势草食性动物。化石证据显示泰坦巨龙类取代其他的蜥脚类恐龙,例如梁龙科与腕龙科,这群动物在晚侏罗纪到中白垩纪期间逐渐被取代、灭亡。
泰坦巨龙类的分布广泛,尤其是南方各大陆(冈瓦纳大陆),甚至在澳大利亚昆士兰州也发现属于泰坦巨龙类的蜥脚类化石。在昆士兰州内地小镇所发现的新标本,身长约为25米长,发现于有9,600万前历史的岩层,显示澳大利亚曾经有大型的泰坦巨龙类生存者。在2006年3月2日有报道传出,在意大利发现了一种泰坦巨龙类的四个保存良好的骨骸。在2009年,新西兰也发现疑似泰坦巨龙类的化石。目前只有南极洲曾没发现过泰坦巨龙类的化石。
分类泰坦巨龙类是群分布广泛而且成功的演化支,但它们的化石纪录非常少。泰坦巨龙类大约有50个种,但直到最近才发现了头颅骨与相对完整的骨骸,例如掠食龙。许多物种的所知有限,随着对于这个演化支的了解增加,许多的化石材料被视为无效,或被重新分类。
泰坦巨龙科是以所知有限的泰坦巨龙属为名,泰坦巨龙属是在1877年由理查德·莱德克(Richard Lydekker)所建立,根据一个部份股骨与两个不完整背椎。曾经有14个种被归类于泰坦巨龙属,使泰坦巨龙属分布于阿根廷、欧洲、马达加斯加、印度、以及老挝,并横跨白垩纪约6,000万年。尽管泰坦巨龙属于泰坦巨龙类的分类系统与生物地理学,但对于泰坦巨龙属所有种的重新审视,使泰坦巨龙属只剩下5个已承认种。模式种印度泰坦巨龙(T. indicus)是无效种,因为该种原本是根据将废弃的特征。因此,泰坦巨龙属的使用大量地遭到遗弃。泰坦巨龙属中的最著名标本从此被重新归类于其他属,包含伊希斯龙。然而,如果印度泰坦巨龙在未来被重新叙述,根据新的发现,泰坦巨龙属将暂时性变成有效属。
有些古生物学家,例如保罗·塞里诺(Paul Sereno)的2005年定义,提出泰坦巨龙属因为资讯过少,因此不能作为定义的基准点,所以不能将其他属归类于以泰坦巨龙属为模式属的科(例如泰坦巨龙超科、泰坦巨龙科、泰坦巨龙亚科)。Weishampel等人在《The Dinosauria》一书的第二版,也没有使用泰坦巨龙科,而是使用数个范围较小的科,例如萨尔塔龙科与纳摩盖吐龙科。
分类学
以下主要根据汤玛斯·荷兹(Thomas Holtz)等人的2012年研究:
泰坦巨龙类 Titanosauria
埃拉尔巨龙 Elaltitan
纳拉姆布埃纳巨龙 Narambuenatitan
南极龙科Antarctosauridae
纳摩盖吐龙科Nemegtosauridae
萨尔塔龙科Saltasauridae
银龙科Argyrosauridae
风神龙科Aeolosauridae
隆柯龙类Lognkosauria
岩盔龙类Lithostrotia
阿马格巨龙 Amargatitanis (阿根廷)
安第斯龙 Andesaurus (阿根廷)
Atacamatitan (智利)
吉普赛龙 Atsinganosaurus (法国)
澳洲南方龙 Austrosaurus (澳洲)
俾路支龙 Balochisaurus (巴基斯坦)
巴罗莎龙 Barrosasaurus (阿根廷)
包鲁巨龙 Baurutitan (巴西)
布罗希龙 Brohisaurus (巴基斯坦)
似倾齿龙 Campylodoniscus (阿根廷)
丘布特龙 Chubutisaurus (阿根廷)
戈壁巨龙 Gobititan (中国)
高桥龙 Hypselosaurus (罗马尼亚)
朱特龙 Iuticosaurus (英格兰)
江山龙 Jiangshanosaurus (中国)
九台龙 Jiutaisaurus (中国)
卡龙加龙 Karongasaurus (马拉维)
海特兰龙 Khetranisaurus (巴基斯坦)
拉布拉达龙 Laplatasaurus (阿根廷、乌拉圭)
利加布龙 Ligabuesaurus (阿根廷)
大尾龙 Macrurosaurus (英格兰)
马拉圭龙 Malarguesaurus (阿根廷)
Marisaurus (巴基斯坦)
巴基龙 Pakisaurus (巴基斯坦)
沼泽巨龙 Paludititan (罗马尼亚)
普尔塔龙 Puertasaurus (阿根廷)
清秀龙 Qingxiusaurus (中国)
汝阳龙 Ruyangosaurus (中国)
苏莱曼龙 Sulaimanisaurus (巴基斯坦)
怪味龙 Tangvayosaurus (老挝)
泰坦巨龙 Titanosaurus (印度)
乌贝拉巴巨龙 Uberabatitan (巴西)
温顿巨龙 Wintonotitan (澳洲)
真泰坦龙类 Eutitanosauria
科学分类
安第斯龙科Andesauridae科
萨尔塔龙科 Saltasauridae
纳摩盖吐龙科 Nemegtosauridae
南极龙科Antarctosauridae
本词条内容贡献者为:
张岩 - 副研究员 - 北京大学分子医学研究所
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国