

简介
三叠纪时大多数地球上的大陆形成一块巨大的大陆:盘古大陆。
三叠纪的名称是1834年弗里德里希·冯·阿尔伯提起的,他将在中欧普遍存在的位于白色的石灰岩和黑色的页岩以及其间的红色的三层岩石层统称为三叠纪。今天,三叠纪被分成更多亚层。三叠纪时的气候炎热干燥,这形成了现在可以看到的当时留下来的典型的红色沙石。当时季节分明,有强烈的雨季。在两极比较潮湿温和。
标志三叠纪的典型的红色沙岩说明当时的气候比较温暖干燥,没有任何冰川的迹象。今天一般认为当时在两极没有陆地或覆冰。因为当时地球上只有一个大陆,因此当时的海岸线比今天要短得多,
时遗留下来的近海沉积比较少,只有在西欧比较丰富。因此三叠纪的分层主要是依靠暗礁地带的生物化石来分的。
由于三叠纪以一次灭绝事件开始,因此其生物开始时分化很厉害。六放珊瑚亚纲是这时候出现的,第一批被子植物和第一种会飞的脊椎动物(翼龙)可能也是这时候出现的。世界上最早的乌龟——原颚龟也出现在三叠纪晚期。第一批鱼龙出现了。
发源历史三叠纪的开始和结束各以一次灭绝事件为标志。虽然这段时间的岩石标志非常明显和清晰,其开始和结束的准确时间却如同其它古远的地质时代无法非常精确地被确定,其误差在正负数百万年。
三叠纪的名称是1834年弗里德里希·冯·阿尔伯提起的,他将在中欧普遍存在的位于白色的石灰岩和黑色的页岩之间的红色的三层岩石层统称为三叠纪。今天,三叠纪被分成更多亚层。
标志三叠纪的典型的红色沙岩说明当时的气候比较温暖干燥,没有任何冰川的迹象。今天一般认为当时在两极没有陆地或覆冰。因为当时地球上 只有一个大陆,因此当时的海岸线比今天要短得多,三叠纪时遗留下来的近海沉积比较少,只有在西欧比较丰富。因此三叠纪的分层主要是依靠暗礁地带的生物化石来分的。
只有一个大陆,因此当时的海岸线比今天要短得多,三叠纪时遗留下来的近海沉积比较少,只有在西欧比较丰富。因此三叠纪的分层主要是依靠暗礁地带的生物化石来分的。
由于三叠纪以一次灭绝事件开始,因此其生物开始时分化很厉害。六放珊瑚亚纲是这时候出现的,第一批被子植物和第一种会飞的脊椎动物(翼龙)可能也是这时候出现的。
三叠纪时的气候炎热干燥,这形成了现在可以看到的当时留下来的典型的红色沙石。当时季节分明,有强烈的雨季。在两极比较潮湿温和。
详细资料名字来源日本首先将希腊文“Trias”译为三叠纪,我国地质界沿用了这一名称。 此期形成的地层称为三叠系,代表符号为“T”。三叠纪分为早、中、晚三个世。
此期形成的地层称为三叠系,代表符号为“T”。三叠纪分为早、中、晚三个世。
生物变革方面,陆生爬行动物比二叠纪有了明显的发展。古老类型的代表(如无孔亚纲和下孔亚纲)基本绝灭,新类型大量出现,并有一部分转移到海中生活。原始哺乳动物在三叠纪末期也出现了。由于陆地面积的扩大,淡水无脊椎动物发展很快,海生无脊椎动物的面貌也为之一新。菊石、双壳类、有孔虫成为划分与对比地层的重要门类,而筳及四射珊瑚则完全绝灭。
爬行动物在三叠纪崛起,主要由槽齿类、恐龙类、似哺乳的爬行类组成。典型的早期槽齿类表现出许多原始的特点,且仅限于三叠纪,其总体结构是后来主要的爬行动物以至于鸟类的祖先模式;恐龙类最早出现于晚三叠世,有两个主要类型:较古老的蜥臀类和较进化的鸟臀类。海生爬行类在三叠纪首次出现,由于适应水中生活,其体形呈流线式,四肢也变成桨形的鳍;似哺乳爬行动物亦称兽孔类,四肢向腹面移动,因此更适于陆地行走。
原始的哺乳动物最早见于晚三叠世,属始兽类,所见到的化石都是牙齿和颌骨的碎片。三叠纪时,晚二叠世幸存的齿菊石类大量繁盛起来,中、晚三叠世的大部分菊石有发达的纹饰,有许多科是三叠纪所特有的。菊石的迅速演化为划分和对比地层创造了极重要的条件。
双壳类也有明显变化,晚古生代的种类只有很少数继续存在,产生了许多新种类,并且数量相当繁多。尤其在晚三叠世,一些种属的结构类型变得复杂,个体也往往比较大。由于三叠纪的环境与古生代不同,非海相双壳类逐渐繁盛起来。
裸子植物的苏铁、本内苏铁、尼尔桑、银杏及松柏类的植物自三叠纪起迅速发展起来。其中除本内苏铁目始于三叠纪外,其它各类植物均在晚古生代就始有了发展,但并不占重要地位。二叠纪的干燥性气候延续到了早、中三叠世,到了中三叠世晚期植物才开始逐渐繁盛。晚三叠世时,裸子植物真正成了大陆植物的主要统治者。1
气候代表三叠纪的典型红色砂岩向我们表明,当时的气候比较炎热干燥,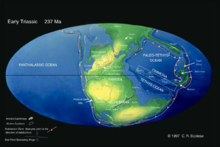 没有任何冰川的迹象,那时的地球两极并没有陆地或覆冰。地球表面的地理分布决定了各地的气候,靠近海洋的地方自然是比较湿润而草木茂盛,但是由于陆地的面积十分广阔,使带湿气的海风无法进入内陆地区,大陆中部便形成了一个很大的沙漠,所以陆地上的气候相当干燥,这进而使得较耐旱的蕨类品种及不过分依赖水繁殖的针叶树逐渐在这些地区取得了竞争优势。
没有任何冰川的迹象,那时的地球两极并没有陆地或覆冰。地球表面的地理分布决定了各地的气候,靠近海洋的地方自然是比较湿润而草木茂盛,但是由于陆地的面积十分广阔,使带湿气的海风无法进入内陆地区,大陆中部便形成了一个很大的沙漠,所以陆地上的气候相当干燥,这进而使得较耐旱的蕨类品种及不过分依赖水繁殖的针叶树逐渐在这些地区取得了竞争优势。
陆地三叠纪的陆地三叠纪时期的地球与现今的地球截然不同,只有一 块大陆,这块大陆被称为泛古陆,即大冈瓦纳古陆。
块大陆,这块大陆被称为泛古陆,即大冈瓦纳古陆。
劳拉西亚古陆包括了今日的北美洲、欧洲和亚洲的大部分地区,冈瓦纳古陆则包括了非洲、大洋州、南极洲、南美洲以及亚洲的印度等部分地区。
不过到三叠纪中期,泛古陆开始出现分裂的前兆,在北美洲、欧洲中部和西部、非洲的西北部均出现了裂痕。
海洋泛古陆之外的地表上是一片一望无际的超大海洋,这个海洋横跨两万多千米,面积大小和今天的所有海洋的总面积差不多。而且由于当时地球上只有一个大陆,因此当时的海岸线比今天要短得多。三叠纪时遗留下来的近海沉积比较少,并且大多分布在如今的西欧地区,因此三叠纪的分层主要是依靠暗礁地带的生物化石来确定的。
地层世界的三叠系广泛分布于、和的边缘海域、及各类拼贴上,也见于大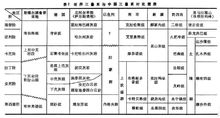 陆内部的内陆盆地内。按照沉积相特点,国际上通常分为海陆交互相的德国型三叠系、海相的阿尔卑斯型三叠系和陆相红层的英国型三叠系。有的学者又提出特提斯南缘浅海相的塞伐狄克型三叠系和北方海域的北极型三叠系等。这些类型实际上侧重于反映欧洲地区的一般情况。此外,还有亚澳地区的陆相含煤三叠系等类型。
陆内部的内陆盆地内。按照沉积相特点,国际上通常分为海陆交互相的德国型三叠系、海相的阿尔卑斯型三叠系和陆相红层的英国型三叠系。有的学者又提出特提斯南缘浅海相的塞伐狄克型三叠系和北方海域的北极型三叠系等。这些类型实际上侧重于反映欧洲地区的一般情况。此外,还有亚澳地区的陆相含煤三叠系等类型。
世界三叠纪地层对比见表1(世界三叠系与中国三叠系对比简表)
生物年代地层分类和对比 18世纪末叶至19世纪中叶,世界三叠系的生物年代分类标准,逐步地在东阿尔卑斯地区建立起来。这就是在奥地利和意大利境内的下三叠统维尔芬阶或斯西提阶,中三叠统安尼阶、拉丁阶、上三叠统卡尼阶、诺利阶和瑞替阶。这些阶的层型,在奥地利的梯罗尔州维尔芬附近,恩斯河流域和意大利的多洛麦特山等地。20世纪以来,三叠系的生物年代分类研究更为详细。主要涉及下三叠统的分阶,二叠-三叠系界线,瑞替阶的归属和各阶的含义厘定等。经过深入研究,德国型与阿尔卑斯型三叠系的生物年代对比也比较明确。2
地层分类对比见表2(三叠系的阶、亚阶和带)
沿革裸子植物的兴盛时代三叠纪是中生代的第一个纪,是古生代生物群消亡后现代生物群开始 形成的过渡时期。海洋无脊椎动物类群发生了重大变化,内生、游泳的软体动物—甲壳动物群落取代表生、固着的腕足动物—海百合群落而成为海洋中的优势群落;六射珊瑚取代四射珊瑚,并迅速发展,遍及全球。与古生代相比,双壳类和菊石类也多属新发展的种类,菊石多具有复杂的纹饰和菊石式缝合线。
形成的过渡时期。海洋无脊椎动物类群发生了重大变化,内生、游泳的软体动物—甲壳动物群落取代表生、固着的腕足动物—海百合群落而成为海洋中的优势群落;六射珊瑚取代四射珊瑚,并迅速发展,遍及全球。与古生代相比,双壳类和菊石类也多属新发展的种类,菊石多具有复杂的纹饰和菊石式缝合线。
三叠纪时,脊椎动物得到了进一步的发展。其中,槽齿类爬行动物出现,并从它发展出最早的恐龙,三叠纪晚期,蜥臀目和鸟臀目都已有不少种类,恐龙已经是种类繁多的一个类群了,在生态系统占据了重要地位因此,三叠纪也被称为“恐龙世代前的黎明”。与此同时,从兽孔类爬行动物中演化出了最早的哺乳动物—似哺乳爬行动物,但是,在随后从侏罗纪到白垩纪长达1亿多年的漫长岁月里,这批生不逢时哺乳动物一直生活在以恐龙为主的爬行动物的阴影之下,直到新生代才成为地球的主宰。
三叠纪早期植物面貌多为一些耐旱的类型,随着气候由半干热、干热向温湿转变,植物趋向繁茂,低丘缓坡则分布有和现代相似的常绿树,如松、苏铁等,而盛产于古生代的主要植物群几乎全部灭绝。3
生物变革灭绝事件
三叠纪以一次灭绝事件结束,尤其对海洋生物来说它的摧毁惨重:牙形石灭绝,除鱼龙外所有的海生爬行动物消失。腕足动物、腹足动物和贝壳等无脊椎动物受到巨大冲击。在海洋中,22%的属,大约一半的种消失。
这次灭绝事件并非在所有地方的摧残程度都一样。在有些地方几乎它没有任何影响。在其它一些地方实际上所有的迷龙和大多数合弓类动物都消失了。许多早的恐龙也均灭绝,而那些发达一些的恐龙却幸存了。许多槽齿目动物也都灭绝了。幸存的植物包括针叶类和苏铁。
这次灭绝事件的原因还不清楚。在2.08至2.13亿年以前盘古大陆开始分裂,这导致了强烈的火山运动,这是地球大陆形成后最强烈的火山运动了。其它可能的原因有全球性的气候冷却或陨星。加拿大魁北克的一处陨石坑曾一度被认作是这次灭绝事件的起因,但后来的调查认为这个陨石坑是在2.14亿年前形成的,比三叠纪的结束早了120±20万年,因此不太可能是这次灭绝事件的直接原因。
甚至连灭绝的确切时间也不十分确定。一些研究认为实际上当时有两次灭绝事件,其相隔时间是1200至1700万年。 这次灭绝事件为恐龙的发展提供了巨大的机会。恐龙在此后的1.5亿年中是地球上最主要、种类最多和数量最大的动物群。

 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国