

转运RNA(Transfer RNA),又称传送核糖核酸、转移核糖核酸,通常简称为tRNA,是一种由76-90个核苷酸所组成的RNA,其3'端可以在氨酰-tRNA合成酶催化之下,接附特定种类的氨基酸。转译的过程中,tRNA可借由自身的反密码子识别mRNA上的密码子,将该密码子对应的氨基酸转运至核糖体合成中的多肽链上。每个tRNA分子理论上只能与一种氨基酸接附,但是遗传密码有简并性(degeneracy),使得有多于一个以上的tRNA可以跟一种氨基酸接附。
研究历史在tRNA被发现以前,佛朗西斯·克里克就假设有种可以将RNA讯息转换成蛋白质讯息的适配分子存在。1960年代早期,亚历山大·里奇、唐纳德·卡斯帕尔等生物学家开始研究tRNA的结构,1965年,罗伯特·W·霍利首次分离了tRNA,并阐明了其序列与大致的结构,他因此贡献而获得1968年的诺贝尔生理学或医学奖。tRNA最早由罗伯特·M·博克(Robert M. Bock)成功结晶,之后陆续有人提出tRNA苜蓿叶状的二级结构,此结构于1973年由金成镐与亚历山大·里奇的X射线衍射分析证实。另一个由阿龙·克卢格领导英国团队,在同一年发布同样的射线晶体学的发现。1
定义大多数tRNA由七十几至九十几个核苷酸折叠形成的三叶草形短链组成,相对分子质量为25000〜30000,沉降常数约为4S。旧称联接RNA、可溶性RNA等。主要作用是携带氨基酸进入核糖体,在mRNA指导下合成蛋白质,即以mRNA为模板,将其中具有密码意义的核苷酸顺序翻译成蛋白质中的氨基酸顺序。tRNA与mRNA是通过反密码子与密码子相互作用而发生关系的。在肽链生成过程中,第一个进入核糖体与mRNA起始密码子结合的tRNA称为起始tRNA,其余tRNA参与肽链延伸,称为延伸tRNA,按照mRNA上密码的排列,携带特定氨基酸的tRNA依次进入核糖体。形成肽链后,tRNA即从核糖体释放出来。整个过程称为tRNA循环。一种tRNA只能携带一种氨基酸,如丙氨酸tRNA只携带丙氨酸,但一种氨基酸可被不止一种tRNA携带。同一生物中,携带同一种氨基酸的不同tRNA称作“同功受体tRNA”。组成蛋白质的氨基酸有20种,根据密码子摆动学说至少需要31种tRNA,但在脊椎动物中只存在22种tRNA。这主要是通过密码子-反密码子配对的简化实现的,使得一种tRNA可以识别一个密码子家族的全部4个密码子。携带同一种氨基酸的细胞器tRNA与细胞质tRNA也不一样。生物体发生突变后,校正机制之一是通过校正基因合成一类校正tRNA,以维持翻译作用译码的相对正确性。可以有多种校正tRNA携带同一种氨基酸。
结构 转运RNA分子由一条长70~90个核苷酸并折叠成三叶草形的短链组成的。上图中有两种不同的分子,苯丙氨酸tRNA(4tna)和天冬氨酸tRNA(2tra)。tRNA链的两个末端在图上方指出的L形结构的末端互相接近。氨基酸在箭头示意的位置被连接。在这条链的中央形成了L形臂,如图下方所示,露出了形成反密码子的三个核苷酸。三叶草结构的其余两环被包裹成肘状,在那里它们提供整个分子的结构。四个常见RNA碱基---腺嘌呤,尿嘧啶,鸟嘌呤和胞嘧啶显然不能提供足够的空间以形成一个坚固的结构,因为这些碱基大部分被修饰过以延长它们的结构。有两个奇特的例子,看37号反密码子相邻的碱基,位于甲硫氨酸tRNA(1yfg)或苯丙氨酸tRNA(4tna和6tna)的起始部位。
转运RNA分子由一条长70~90个核苷酸并折叠成三叶草形的短链组成的。上图中有两种不同的分子,苯丙氨酸tRNA(4tna)和天冬氨酸tRNA(2tra)。tRNA链的两个末端在图上方指出的L形结构的末端互相接近。氨基酸在箭头示意的位置被连接。在这条链的中央形成了L形臂,如图下方所示,露出了形成反密码子的三个核苷酸。三叶草结构的其余两环被包裹成肘状,在那里它们提供整个分子的结构。四个常见RNA碱基---腺嘌呤,尿嘧啶,鸟嘌呤和胞嘧啶显然不能提供足够的空间以形成一个坚固的结构,因为这些碱基大部分被修饰过以延长它们的结构。有两个奇特的例子,看37号反密码子相邻的碱基,位于甲硫氨酸tRNA(1yfg)或苯丙氨酸tRNA(4tna和6tna)的起始部位。
一级结构
自1965年R.W.霍利等首次测出酵母丙氨酸tRNA的一级结构即核苷酸排列顺序到1983年已有200多个tRNA(包括不同生物来源、不同器官、细胞器的同功受体tRNA以及校正tRNA)的一级结构被阐明。按照A-U、G-C以及G-U碱基配对原则,除个别例外,
二级结构
tRNA分子均可排布成三叶草模型的二级结构(图1)。它由3个环,即D环〔因该处二氢尿苷酸(D)含量高〕、反密码环(该环中部为反密码子)和TΨC环〔因绝大多数tRNA在该处含胸苷酸(T)、假尿苷酸(Ψ)、胞苷酸(C)顺序〕,四个茎,即D茎(与D环联接的茎)、反密码茎(与反密码环联接)、TΨC茎(与 TΨC环联接)和氨基酸接受茎〔也叫CCA茎,因所有tRNA的分子末端均含胞苷酸(C)、胞苷酸(C)、腺苷酸(A)顺序, CCA是连接氨基酸所不可缺少的〕,以及位于反密码茎与TΨC茎之间的可变臂构成。不同tRNA的可变臂长短不一,核苷酸数从二至十几不等。除可变臂和D环外,其他各个部位的核苷酸数目和碱基对基本上是恒定的。图1也示出tRNA分子中出现的保守或半保守成分。这些成分对维系tRNA的三级结构是很重要的。
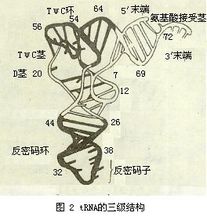
tRNA的结构特征
tRNA的结构特征之一是含有较多的修饰成分,如上面提到的 D、T、 Ψ等;核酸中大部分修饰成分是在tRNA中发现的。修饰成分在tRNA分子中的分布是有规律的,但其功能不清楚。
1974年用X射线晶体衍射法测出第一个tRNA——酵母苯丙氨酸tRNA晶体的三维结构,分子全貌象倒写的英文字母L,呈扁平状,长60埃,厚20埃(图2),它是在tRNA二级结构基础上,通过氨基酸接受茎与TΨC茎以及D茎与反密码茎间折叠成右手反平行双螺旋。tRNA三级结构由保守或半保守成分与构成二级结构的核苷酸之间形成氢键(称三级结构氢键)维系。其他tRNA晶体的三维结构类似酵母苯丙氨酸tRNA,只是某些参数有所不同。tRNA在溶液中的构型与其晶体结构一致。
功能主要是携带氨基酸进入核糖体,在mRNA指导下合成蛋白质。即以mRNA为模板,将其中具有密码意义的核苷酸顺序翻译成蛋白质中的氨基酸顺序(见蛋白质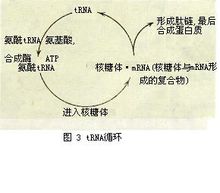 的生物合成、核糖体)。tRNA与mRNA是通过反密码子与密码子相互作用而发生关系的。在肽链生成过程中,第一个进入核糖体与mRNA起始密码子结合的tRNA叫起始tRNA,其余tRNA参与肽链延伸,称为延伸tRNA,按照mRNA上密码的排列,携带特定氨基酸的tRNA依次进入核糖体。形成肽链后,tRNA即从核糖体释放出来。整个过程叫做tRNA循环(图3)。tRNA靠反密码子与mRNA识别,但并非一种反密码子只能识别一种密码子。例如反密码子CIG(I是次黄嘌呤核苷酸)能识别三种密码子。一般反密码子中的稀有核苷酸因配对不严格而能识别多种密码子,这种现象在生物学中称为“摆动性”
的生物合成、核糖体)。tRNA与mRNA是通过反密码子与密码子相互作用而发生关系的。在肽链生成过程中,第一个进入核糖体与mRNA起始密码子结合的tRNA叫起始tRNA,其余tRNA参与肽链延伸,称为延伸tRNA,按照mRNA上密码的排列,携带特定氨基酸的tRNA依次进入核糖体。形成肽链后,tRNA即从核糖体释放出来。整个过程叫做tRNA循环(图3)。tRNA靠反密码子与mRNA识别,但并非一种反密码子只能识别一种密码子。例如反密码子CIG(I是次黄嘌呤核苷酸)能识别三种密码子。一般反密码子中的稀有核苷酸因配对不严格而能识别多种密码子,这种现象在生物学中称为“摆动性”
tRNA是通过分子中3′端的CCA携带氨基酸的。氨基酸连接在腺苷酸的2′或3′OH基上,携带了氨基酸的tRNA叫氨酰tRNA,例如,携带甘氨酸的tRNA叫甘氨酰tRNA。氨基酸与tRNA的结合由氨酰tRNA合成酶催化,分二步进行:①氨基酸+ATP→氨酰-AMP+焦磷酸;②氨酰-AMP+tRNA→氨酰-tRNA+AMP。与一种氨基酸对应的至少有一种tRNA和一种氨酰-tRNA合成酶(见蛋白质生物合成)。
tRNA还具有其他一些特异功能,例如,在没有核糖体或其他核酸分子参与下,携带氨基酸转移至专一的受体分子,以合成细胞膜或细胞壁组分;作为反转录酶引物参与DNA合成;作为某些酶的抑制剂等。有的氨酰-tRNA还能调节氨基酸的生物合成。在许多植物病毒RNA分子中发现有类似于tRNA的三叶草结构,有的也能接受氨基酸,其功能不详2。
合成方法生物合成:在生物体内,DNA分子上的tRNA基因经过转录生成tRNA前体,然后被加工成成熟的tRNA:
tRNA前体的加工包括:切除前体分子中两端或内部的多余核苷酸;形成tRNA成熟分子所具有的修饰核苷酸;如果前体分子3′端缺乏CCA顺序,则需补加上CCA末端。加工过程都是在酶催化下进行的。
人工合成:1981年,中国科学家王德宝等用化学和酶促合成相结合的方法首次全合成了酵母丙氨酸tRNA。它由76个核苷酸组成,其中包括天然分子中的全部修饰成分,产物具与天然分子相似的生物活性(见核糖核酸和核酸人工合成)。
本词条内容贡献者为:
成玉林 - 副教授 - 重庆大学
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国