

鞭毛纲(Mastigophora)指的是原生动物门,质走亚门的一纲。物种约10000种。一般具鞭毛,以鞭毛为运动器,鞭毛通常有1~4条或稍多,鞭毛内周缘部排列9条双联体微管、中央有2条中央微管。无性繁殖一般为纵二分裂,有性繁殖为配子结合或整个个体结合。在环境不良的条件下一般能形成包囊。根据营养方式的不同,可分为两个亚纲:植鞭亚纲Phytomastigina一般具有色素体能行光合作用,自由生活在淡水或海水中(自养),种类多,形状各异,如眼虫Euglena、盘藻Gonium、团藻Volvox、夜光虫Noctiluca(腰鞭毛虫类,可引发赤潮)、钟罩虫Dinobryon、尾窝虫Uroglena、合尾滴虫Synura。动鞭亚纲Zoomastigina无色素体,寄生、腐生或吞噬营养(异养),如利什曼原虫Leishmania(黑热病,白蛉子传播),锥虫Trypanosoma(昏睡病)、隐鞭虫Cryptobia(寄生于鱼鳃)、披发虫Ttrichonympha(共生于白蚁肠道,使其可以消化木材的纤维素)。
概述鞭毛纲 原生动物门的一纲。以鞭毛运动。一般具有104根鞭毛,少数种类具有较多的鞭毛。有些种类具有色素体,能进行光合作用,自己制造食物,为植物性营养方式。有的通过体表渗透吸收周围呈溶解状态的物质,为腐生性营养。另一些则吞食固体的食物颗粒,为动物性营养方式。其中植物性营养方式为本纲所特有。有的种类根据生活环境条件的改变,或兼有两种营养方式。在自然情况下都以二分裂法进行繁殖,除此,也有能形成配子进行有性生殖的。鞭毛虫有自由生活于淡水或海水中的(眼虫、夜光虫等),也有营寄生生活的。有不少寄生种类,能引起人或家畜的疾病(如引起黑热病的利什曼原虫,寄生于人或家畜体内的锥虫、寄生于鱼鳃的鳃隐鞭虫等)。鞭毛纲还有些种类以致个或许多个体相聚,成为群体的(盘藻、团藻等)1。
简介鞭毛纲(Mastigophora)动物在其成年阶段都有一根、几根或许多根鞭毛作为其运动细胞器,由于它们营养方式及生殖方式的原始性和多样性,一般认为鞭毛虫纲是最原始的原生动物,通过它可以将动物界与植物界联系起来。鞭毛虫纲可以分为两个亚纲:植鞭毛亚纲(Phytomastigina)及动鞭毛亚纲(Zoomastigina),前者都具有色素体,可行光合作用即进行植物性营养。淀粉及副淀粉是其主要的食物贮存物,有无性生殖和有性生殖,虫体通常具有2根鞭毛,体表为皮膜或纤维素细胞壁,自由生活。后者没有色素体,行动物性营养或腐生性营养,糖元是其食物贮存物,不行有性生殖,有一到数根鞭毛,虫体表面只有细胞膜,除少数种类自由生活外,多数种类在多细胞动物体内营共生或寄生生活。
主要特征原生动物门的一种,已知的有2000个种类。其主要特征为:
1.鞭毛。为本纲动物最主要的特征·鞭毛不仅是运动胞器,而且还具有感觉及帮助摄食作用.不同种类的鞭毛数目和着生方式等不同。
2.营养方式t因种类不同,鞭毛纲具有原生动物三种营养类型,即植物性营养(像植物一样利用阳光合成有机物)、动物性营养(吞食其他生物或有机碎片)和渗透性营养(靠体表渗透摄取有机物质)。
3.生殖:无性生殖为二分纵裂,如绿眼虫。夜光虫能进行出芽生殖。有性生殖为同配生殖,如衣藻虫}异配生殖,如困藻虫等2。
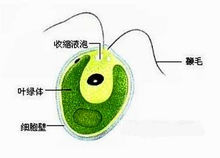
基本形态鞭毛虫类身体呈卵圆形、球形、长圆形等,通常能分出前后端,都以鞭毛作为运动器官,体表具细胞膜、或皮膜或细胞壁。例如眼虫(Euglena),体表具有皮膜(或称表膜),是由一层蛋白质组成螺旋形排列的条纹,在条纹上有嵴和沟,使皮膜呈现斜纹,眼虫的皮膜很薄、富有弹性,使身体可以收缩改变形状,做眼虫式运动(euglenoid movement)。鞭毛虫缺乏明显的内质与外质的分化。眼虫身体的前端有一凹陷的沟,称储蓄泡(reservoir)或称胞咽,但它不是取食的器官,是水的贮存处及鞭毛着生部位,基体位于它的基部,由基体伸出两根鞭毛,一根很长伸向体外,一根很短不伸出储蓄泡。眼虫和其他一些植鞭毛虫类在身体的前端含有类胡萝卜素(carotinoid)的脂类球集合而成的红色眼点,这种色素球只有在基质中含有很少的氮及磷时才出现,否则将消失,它可以吸收光线。在鞭毛的基部具有副鞭毛体(paraflagellar body),是一种光敏感结构,当一侧的光被眼点吸收后,可使它产生一个定向的光反应,所以副鞭毛体与眼点构成了某些植鞭毛虫类的感光细胞器。眼虫和许多淡水鞭毛虫有一个或多个伸缩泡。眼虫的伸缩泡位于身体前端,它是由几个小泡收集了细胞质内过多的水分愈合而成,当伸缩泡充满水分之后,排到储蓄泡中,再通过其开口排到体外。某些鞭毛虫没有储蓄泡,则伸缩泡内的水分通过体表排到体外。海洋或寄生的种类伸缩泡不发达或完全不存在。眼虫及其他植鞭毛虫类,细胞内散布有形状、数目不等的色素体,眼虫的色素体中含有叶绿素a和b,胡萝卜素(carotin)和叶黄素,使虫体也表现出绿色。有些植鞭毛虫的叶绿素被其他色素所隐蔽,而使鞭毛虫表现出黄绿色或黄褐色等,与色素体常伴随出现的是淀粉核(pyrenoid),它对副淀粉体的形成起作用。大多数鞭毛虫具有一个大的细胞核,位于身体的近后端,少数种类具有2个核,核内有核内质(endosome)。眼虫的细胞核在分裂间期时染色质浓缩,分裂过程中不行成纺锤体,核膜不消失,这是很原始的现象。
鞭毛与运动鞭毛虫纲是以鞭毛作为运动器官,当鞭毛虫有两根或许多根鞭毛时,它们等长或不等长,如果不等长,其中一根起主导作用,另一根是附属的,例如囊杆虫(Peranema)。眼虫在运动时也是一根鞭毛起作用。许多植鞭毛虫的鞭毛可向一侧或两侧伸出许多平行排列的细丝,称茸鞭茸毛(mastigonemes),具有两根不等长鞭毛的种类,一般主鞭毛是茸鞭茸毛,它的作用可能是逆转水的推动力。一些寄生的鞭毛虫其鞭毛不是游离的,它由基体发生之后,与身体表面的细胞质膜之间有一原生质相连,在电子显微镜下观察,是成行密集排列的丝状结构连接在鞭毛与细胞之间,鞭毛的末端,离开身体之后是游离的,例如锥虫,这样的结构有利于鞭毛虫在粘滞性很高的血液或体液中运动。鞭毛运动的机制尚不十分清楚,不同的种类其鞭毛使用的方法并不完全相同。早期的理论认为鞭毛在运动时是螺旋旋转,像推进器一样推动身体前进。知道鞭毛运动更多的是划动(rowing)及波动(undulating)。划动是鞭毛与附着点成直角方向在水中甩动,随之是鞭毛的恢复打动。借助于鞭毛甩动时水流的反作用力推动身体前进,并沿身体的纵轴向前旋转前进,或左右摆动前进。波动运动是鞭毛在一个平面上呈波状摆动。如果波状摆动是由鞭毛的基部向端部发生,则驱使身体向相反的方向运动,即推动身体向后运动。如果波动是由鞭毛的端部向基部进行,则推动身体向前运动,在有两个鞭毛的种类,运动的方向及路线是由主鞭毛决定的。关于鞭毛的结构与运动机制、人们普遍认为不是由于鞭毛中微管纤维的收缩,因为它们在运动中并不改变长度,而是由成双联体的两个亚纤维彼此滑动,引起鞭毛的弯曲,而亚纤维的滑动也像肌纤维的微丝滑动一样,并且也是利用三磷酸腺苷(ATP)为能源。
营养鞭毛虫类除了没有化学合成营养物之外,可以行各种营养方式。植鞭毛虫类均行植物性营养,也称自养(autotrophs);动鞭毛虫类行动物性或腐生性营养,或两者兼行;统称为异养(heterotrophs)。实际上对许多植鞭毛虫并不能严格地区分自养与异养。例如眼虫在光照条件下,它能自行合成有机物,如把眼虫放在黑暗中饲养一段时间之后,眼虫的叶绿素会逐渐消失,而改行腐生性营养,如果把眼虫再放回光照条件下,它又可重新出现叶绿素,又恢复自养。一些腰鞭毛虫类及金滴虫类可以兼行自养与异养。有一种棕鞭毛虫(Ochromonas)甚至可以兼行自养、动物性营养及腐生性营养三种营养方式。严格的异养仅出现在动鞭毛亚纲,它们或进行吞噬,或在鞭毛的基部形成胞咽取食。营养实验研究表明,鞭毛虫类比其他的原生动物的营养需求更简单。植鞭毛虫类生活的环境中只要有适当伪氮源及碳源,它就可以合成所需要的蛋白质;腐生性的鞭毛虫要求环境中有氨盐,就可以合成蛋白质;在营动物性营养的鞭毛虫则要求环境中有氨基酸及蛋白胨以合成所需要的蛋白质。其他纲原生动物则需要现成的食物(蛋白质),其自行合成食物的生理机能已经消失。植鞭毛虫类食物的贮存形式是植物淀粉,副淀粉体、脂肪及麦清蛋白(leucosin),动鞭毛虫类食物的贮存形式是糖元。
生殖鞭毛虫类的无性生殖主要是二分裂,其中多数又是纵二分裂,例如眼虫,细胞分裂由中心粒(centrioles)或基体的分裂开始,由每个中心粒产生一个新的基体及一根鞭毛,然后每一个新鞭毛与一个老鞭毛根部愈合,同时核也进行有丝分裂,但核膜不消失、不行成纺锤体,随后细胞质由前向后分裂,并复制新的细胞器,最后形成两个相似的子细胞。除了纵分裂,有些种类生殖时可以是:斜分裂或横裂,例如角藻;或多分裂,例如某些腰鞭毛虫类及锥虫等;也可以出现出芽生殖,例如夜光虫(Noctiluca),即由母体上可以长出许多小的芽体。眼虫分裂时一个子细胞保留旧鞭毛,一个子细胞再复制一个新鞭毛,但也有的种原来的鞭毛完全消失,两个子细胞均产生新的鞭毛;如果虫体有几根鞭毛,分裂前部分鞭毛已经复制,分裂时再均等的分配在两个子细胞中。在多鞭毛的种,分裂时鞭毛均等的分配在两个子细胞中,然后每个子细胞再复制一定数量的鞭毛使之恢复到应有的鞭毛数。腰鞭毛虫类体表常具有甲板(armored),分裂时甲板或消失,或均等或不均等的分配到子细胞中。角藻常进行不完全的分裂,结果使多次分裂的子体连成一个长链。群体生活的种类如团藻(Volvox)分裂的结果在群体中形成子群体。总之,无性生殖的方式因种而不同。有性生殖主要出现在植鞭毛虫类及复杂的多鞭毛虫类。它们或是同配生殖或异配生殖,例如衣滴虫(Chlamydomonas)生殖时两个虫体彼此融合,所以虫体本身又是配子,减数分裂发生在配子融合之后,衣滴虫是同配生殖的典型例子。实球虫(Pandorina)是异配生殖,融合的两个配子仅略有大小的区别,大配子仍有鞭毛。团藻是以典型的异配生殖形成精子与卵子,其精子与卵子可能来自同一个群体(雌雄同群体),也可能来自不同的群体(雌雄异群体),卵直接来自群体的生殖细胞其体积增大、没有鞭毛、不会运动。精子是由生殖细胞经多次分裂形成,精子成束、具鞭毛、能游动。卵受精之后,外面可分泌厚壁形成卵囊以越冬。鞭毛虫类的包囊形成是很普遍的,特别是营动物性营养的种类、具叶绿素的种类较少发现包囊。
分类植鞭毛亚纲 Phytomastigina
金滴虫目 Chrysomonadid
隐滴虫目Cryptomonadida
腰鞭毛目 Dinoflagellida
眼虫目 Euglenida
绿滴虫目 Chloromonadida
团藻目 Volvocida
动鞭毛亚纲 Zoomastigina
领鞭毛目 Choanoflagellida
根足鞭毛虫目Rhizomastigida
动质体目 Kinetoplastida
曲滴虫目 Retortamonadida
双滴虫目 Diplomonadida
毛滴虫目 Trichomonadida
超鞭毛目 Hypermastigida3
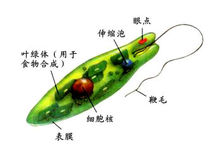
代表性动物眼虫眼虫是眼虫目Euglenida眼虫属Euglena生物的统称,在植物学中称裸藻,是一类介于动物和植物之间的单细胞真核生物。淡水中习见的眼虫有:绿眼虫Euglena viridis,体纺锤形,前端钝圆,后端宽,末端尖呈尾状。鞭毛与体等长,色素体1个,星状。梭眼虫Euglena acus,长纺锤形,鞭毛短,色素体多个。长眼虫Euglena deses,体圆柱形,狭长,鞭毛约为体长的1/3~1/2。螺纹眼虫Euglena spirogyra,体易变形,体表螺旋形带纹明显,鞭毛短。扁眼虫Phacus,体呈宽卵圆形,背腹扁,后端尖刺状,鞭毛与体等长。眼虫生活在有机物质丰富的水沟、池沼或缓流中。但在河堤、海湾湿土或含盐沼泽中亦有之,此外在其它藻类体上、植物碎片、及小甲壳类的体上亦能见到。至于营有机性的种类则多见之于下水道的水内。温暖季节可大量繁殖常使水呈绿色。眼虫体呈绿色梭形,长约60μm,前端钝圆,后端尖。在虫体中部稍后有一个大而圆的核,生活时是透明的。体表覆以具弹性的、带斜纹的表膜。经电子显微镜研究,表膜即质膜或称三分质膜。表膜是由许多螺旋状的条纹联结而成,每一个表膜条纹的一边有向内的沟(groove),另一边有向外的嵴(crest)。一个条纹的沟与其邻接条纹的嵴相关联(似关节)。眼虫生活时,表膜条纹彼此相对移动,可能是由于嵴在沟中滑动的结果。表膜下的粘液体(mucusbody)外包以膜,与体表膜相连续,有粘液管通到嵴和沟。粘液对沟嵴联结的“关节”可能有滑润作用。表膜覆盖整个体表、胞咽、储蓄泡、鞭毛等。使眼虫保持一定形状,又能作收缩变形运动。眼虫体前端有一胞口(cytostome).向后连一膨大的储蓄泡(reservoir),从胞口中伸出一条鞭毛(flagellum)。鞭毛是能动的细胞表面的突起。鞭毛下连有2条细的轴丝(axoneme)。每一轴丝在储蓄泡底都和一基体(basal body)相连,由它产生出鞭毛。基体对虫体分裂起着中心粒的作用。从一个基体连一细丝(根丝体rhizoplast)至核,这表明鞭毛受核的控制。鞭毛最外为细胞膜,其内由纵行排列的微管(microtubule)组成。周围有9对联合的微管(双联体doublets), 中央有2个微管。每个双联体上有2个短臂(arms),对着下一个双联体,各双联体有放射辐(radial spokeS)伸向中心。在双联体之间又有具弹性的连丝(links)。微管由微管蛋白(tubulin)组成,微管上的臂是由动力蛋白(dynein)组成,具有ATP酶的活性。实验证明,鞭毛的弯曲,是由于双联体微合彼此相对滑动的结果,如图所示,在弯曲的内、外侧放射辐的间隔不改变,弯曲是由于弯曲的外侧微管和放射辐对于弯曲内侧的微管和放射辐的相对滑动。一般认为臂能使微管滑动(很像肌肉收缩时,横桥在粗、细肌丝间的滑动),臂上的ATP酶分解ATP提供能量。眼虫借鞭毛的摆动进行运动。眼虫在运动中有趋光性,这是因为在鞭毛基都紧贴着储蓄泡有一红色眼点(stigma),靠近眼点近鞭毛基都有一膨大部分,能接受光线,称光感受器(photoreceptor)。眼点是由埋在无色基质中的类胡萝卜素(carotenoid)的小颗粒组成的。也有人认为是由胡萝卜素(carotene)组成的,或是由β-红萝卜素与血红素组成的。眼点呈浅杯状,光线只能从杯的开口面射到光感受器上,因此,眼虫必须随时调整运动方向,趋向适宜的光线。现在有些学者认为,眼点是吸收光的“遮光物”(light absorbing shade),在眼点处于光源和光感受器之间时,眼点遮住了光感受器,并切断了能量的供应,于是在虫体内又形成另一种调节,使鞭毛打动,调整虫体运动,让光线的连续地照到光感受器上。这样连续调节使眼虫趋向光线。眼点和光感受器普遍存在于绿色鞭毛虫,这与它们进行光合作用的营养方式有关。在眼虫的细胞质内有叶绿体(chloroplast)。叶绿体的形状(如卵圆形、盘状、片状、带状、星状等)、大小、数量及其结构(有无蛋白核及副淀粉鞘)为眼虫属、种的分类特征。眼虫主要通过叶绿素在有光的条件下利用光能进行光合作用,把二氧化碳和水合成糖类,这种营养方式(与一般绿色植物相同),称为光合营养(phototrophy)。制造的过多食物形成一些半透明的副淀粉粒(Paramylum granule)储存在细胞质中。副淀粉粒与淀粉相似,是糖类的一种,但与碘作用不呈蓝紫色.副淀粉粒是眼虫类特征之一,其形状大小也是其分类的依据。在无光的条件下,眼虫也可通过体表吸收溶解于水中的有机物质。这种营养方式称为渗透营养(osmotrophy)。眼虫前端的胞口是否取食固体食物颗粒还有异议。但是已肯定经过胞口可以排出体内过多的水分。在储蓄泡旁边有一个大的伸缩泡(contractile vacuole),它的主要功能是调节水分平衡。收集细胞质中过多的水分(其中也有溶解的代谢废物)。排入储蓄泡,再经胞口排出体外。眼虫在有光的条件下,利用光合作用所放出的氧进行呼吸(氧化)作用,呼吸作用所产生的二氧化碳,又被利用来进行光合作用。在无光的条件下,通过体表吸收水中的氧,排出二氧化碳。眼虫的生殖方法一般是纵二分裂,这也是鞭毛虫纲的特征之一。先是核进行有丝分裂,在分裂时核膜不消失,基体复制为二,继之虫体开始从前端分裂,鞭毛脱去,同时由基体再长出新的鞭毛,或是一个保存原有的鞭毛,另一个产生新的鞭毛。胞口也纵裂为二,然后继续由前向后分裂,断开成为2个个体。在环境不良的条件下。如水池干涸.眼虫体变圆,分泌一种胶质形成包囊,将自己包围起来。刚形成的包囊、可见有眼点,绿色,以后逐流变为黄色,眼点消失,代谢降低,可以生活很久,随风散布于各处。当环境适合时,虫体破囊而出,在出囊前进行一次或几次纵分裂。包囊形成对眼虫度过不良环境是一种很好的适应。多年来用眼虫进行基础理论的研究取得不少成果。不仅对遗传变异理论的探讨有意义,而且对了解有色、无色鞭毛虫类动物间的亲缘关系,对了解动、植物的亲缘关系都有重要意义。近年来也有用眼虫作为有机物污染环境的生物指标,用以确定有机污染的程度,另外眼虫对净化水的放射性物质也有作用。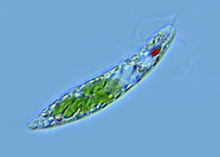
夜光虫夜光虫Noctiluca属腰鞭毛目Dinoflagellida。它们在夜间由于海水波动的刺激能发光,因而得名。夜光虫的身体为圆球形,直径为1毫米左右,颜色发红,细胞质密集于球体的一部分,其内有核,其他部分由细胞质放散成粗网状,在网眼间充满液体。有两根鞭毛,一根较大,称为触手,另一根较小。它的繁殖为分裂法和出芽法两种,后者在身体表面生出很多小的个体,脱离母体后发育成新的个体。例如闪光夜光虫身体的直径为0.5~2毫米,肉眼看到的是一个个晶亮的小球,有透明的细胞膜、网状分散的细胞质、浓密的细胞核、一根细小的鞭毛以及原生质突起形成的粗大可动的触手。因其体内含有许多拟脂颗粒,故受到机械刺激时能发光。在海水中生活的夜光虫和其他一些腰鞭毛虫(如沟腰鞭虫Gonyaulax 、裸甲腰鞭虫Gymnodinium等)大量繁殖,每立方米海水可多达20~40百万个个体,致使海水呈现红色或褐色,称为赤潮,由于它们排出大量代谢产物以及腐败海水,造成沿海鱼类及养殖贝类的大量死亡。腰鞭毛目Dinoflagellida,具两根鞭毛,分别位于身体中部的横沟及后部的纵沟内,横沟内的鞭毛使身体旋转、纵沟内鞭毛推动身体前进。色素体黄色或褐色,少数种无色素体。一些种类体表裸露,如Gymnodinium,多数种类体表有纤维素形成的薄膜、或甲板,如薄甲藻Glenodinium、膝沟藻Gonyaulax等。眼点通常是存在的。无性生殖有的种可行横裂及斜裂,如角藻Ceratium。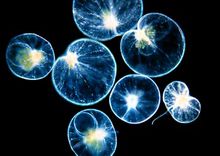
利什曼原虫利什曼原虫Leishmania属动质体目Kinetoplastida(具1~2根鞭毛,所有的成员都有一动质体,常与根丝体相连,其中含有DNA,位于大的延伸的线粒体内,身体一侧有波动膜)。生活史有前鞭毛体(promastigote)和无鞭毛体(amastigote)两个时期。前者寄生于节肢动物(白蛉)的消化道内,后者寄生于哺乳动物或爬行动物的细胞内,通过白蛉传播。对人和哺乳动物致病的利什曼原虫有:引起人体内脏利什曼病的为杜氏利什曼原虫Leishmania donovani,引起皮肤利什曼病的为热带利什曼原虫Leishmania tropica和墨西哥利什曼原虫Leishmania mexicana,引起粘膜皮肤利什曼病的为巴西利什曼原虫Leishmania araziliensis等。我国的黑热病是由杜氏利什曼原虫引起的。杜氏利什曼原虫的无鞭毛体主要寄生在肝、脾、骨髓、淋巴结等器官的巨噬细胞内,常引起全身症状,如发热、肝脾肿大、贫血、鼻衄等。在印度,患者皮肤上常有暗的色素沉着,并有发热,故又称Kala-azar,即黑热的意思。因其致病力较强很少能够自愈,如不治疗常因并发病而死亡。解放前,全国黑热病患者较多。主要流行于长江以北广大地区,为我国五大寄生虫病之一。解放后在党的领导下,在各流行区建立专门的防治机构,发动群众从治病、消灭病犬和白岭子三方面进行防治。现已在全国范围内基本上控制了黑热病的流行。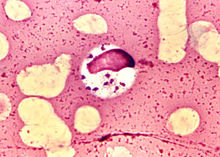
锥体虫锥体虫Trypanosoma动质体目锥虫科锥虫属。体呈柳叶状,运动胞器是一根鞭毛。寄生在血液中的鞭毛虫,其鞭毛与虫体之间连成为波动膜,借以增强在粘滞性较高的血液中的活动能力。细胞核 1个。体内有一称作动基体的特殊细胞器,位于锥虫体内的线粒体内,含有丰富的DNA,谓之动基体DNA(KDNA),能自我复制。动基体的确切功能不明。繁殖方式为二分裂。种类多,分别寄生在各种脊椎动物(鱼类、两栖类、爬行类、鸟类和哺乳类)的血液和组织液中。有个别种类如枯氏锥虫则寄生在人的细胞内。除马媾疫锥虫是通过交媾直接传播外,所有寄生在脊椎动物的锥虫均要依赖某些昆虫(如采采蝇等)进行传播。带有锥虫的脊椎动物血液被媒介动物吸食后,锥虫因生活环境改变,在媒介动物消化道中依次经过无鞭毛体或前鞭毛体上鞭毛体和后循环锥鞭毛体几个不同的发育阶段。在媒介动物中只有这种后循环锥虫对宿主才有感染力。当媒介再次叮咬宿主时,便把后循环锥虫输入宿主体内。伊氏锥虫在媒介消化道中不进行任何发育,它靠媒介在吸血后短时间内作机械传播。锥虫主要通过胞饮作用从宿主获得营养。所有锥虫均可在体外连续培养。锥虫是人和家畜重要的寄生虫之一。按其传播方式可分为两大类:
①粪便型,通过被后循环锥虫污染的粪便传播;
②唾液型,后循环锥虫经唾液腺传播。对人有严重致病作用的锥虫有:罗得西亚锥虫、冈比亚锥虫、枯氏锥虫等。前两者主要流行于非洲各地,引起所谓非洲睡眠病;后者主要分布在南美洲(特别是巴西),引起美洲锥虫病,即夏格氏病。中国还没有发现人体锥虫的病例。对家畜有严重致病作用的锥虫有布氏锥虫、活泼锥虫、刚果锥虫、伊氏锥虫和马媾疫锥虫等。前三者主要流行于非洲,中国尚无记载。伊氏锥虫在中国分布甚广,牛、马伊氏锥虫病即所谓苏拉病,在中国南方各省流行相当严重,造成大批家畜死亡。由于锥虫存在抗原变异的特性,仍未获得有效的疫苗。寄生家畜的布氏锥虫和寄生鼠的路氏锥虫是实验室常用的研究材料。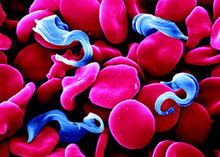
本词条内容贡献者为:
刘敦华 - 教授 - 宁夏大学
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国