

生态位是指一个种群在生态系统中,在时间空间上所占据的位置及其与相关种群之间的功能关系与作用。生态位又称生态龛。表示生态系统中每种生物生存所必需的生境最小阈值。
两个拥有相似功能生态位、但分布于不同地理区域的生物,在一定程度上可称为生态等值生物。生态位的概念已在多方面使用,最常见的是与资源利用谱(resources utilization spectra)概念等同。所谓“生态位宽度”(niche breadth)是指被一个生物所利用的各种不同资源的总和。在没有任何竞争或其它敌害情况下,被利用的整组资源称为“原始”生态位(fundamental niche)。因种间竞争,一种生物不可能利用其全部原始生态位,所占据的只是现实生态位(realized niche)。生态位概念应用于植物尚有一定困难。几乎一切植物都需要阳光进行光合作用,而且在陆地上倾向于占有环境的相同部分——土壤表面上下一定高度和深度。然而,植物种在自然环境水平地带和垂直地带中占据的区域以及在生长季节、开花季节等方面出现的差异,可与动物的生态位相比拟。
定义生态位(ecological niche)是指每个个体或种群在种群或群落中的时空位置及功能关系。
简介生态位又称生态龛。表示生态系统中每种生物生存所必需的生境最小阈值。为食物和空间的竞争,是影响动、植物的种分布的主要生物因子,时常受捕食或植牧的影响。种适应的环境的物理因子限度,大体决定了其地理的或生境的规模的范围,但是种在这一范围内生存,取决于同生活在那里的其它种竞争资源。要了解这对于生物地理学的完整意义,就必须论及生态学理论的一些要点1。
具体介绍生态位又称生态龛。表示生态系统中每种生物生存所必需的生境最小阈值。
内容包含区域范围和生物本身在生态系统中的功能与作用。1924年由格林内尔(J.Gri-nell)首创,并强调其空间概念和区域上的意义。1927年埃尔顿(Charles Elton)将其内涵进一步发展,增加了确定该种生物在其群落中机能作用和地位的内容,并主要强调该生物体对其它种的营养关系。在自然环境里,每一个特定位置都有不同种类的生物,其活动以及与其它生物的关系取决于它的特殊结构、生理和行为,故具有自己的独特生态位。 如每一种生物占有各自的空间,在群落中具有各自的功能和营养位置,以及在温度、湿度、土壤等环境变化梯度中所居的地位。一个种的生态位,是按其食物和生境来确定的。如海星(pisaster ochraceus)在北美洲太平洋沿岸居于主要捕食者的龛位。按竞争排斥原理,任何两个种一般不能处于同一生态龛。在特定生态环境中赢得竞争的胜利者,是能够最有效地利用食物资源和生存空间的种,其种群以出生率高、死亡率低而有较快的增长。有着相似食物或空间要求的数群近缘种,因处不同生态位,彼此并不竞争。麦克阿瑟(R.H. MacArthur)的研究发现,北美洲东北部有5种鸣禽在针叶林里一起生活,都属于林莺属(Den-droica),均以昆虫为食,对营巢地点也有相似要求,但每一种鸣禽在取食和营巢行为上显示了复杂的差别和各自占据的生态位,至少在食物丰富时防止了竞争。
概念应用两个拥有相似功能生态位、但分布于不同地理区域的生物,在一定程度上可称为生态等值生物。生态位的概念已在多方面使用,最常见的是与资源利用谱(resources utilization spectra)概念等同。所谓“生态位宽度”(niche breadth)是指被一个生物所利用的各种不同资源的总和。在没有任何竞争或其它敌害情况下,被利用的整组资源称为“原始”生态位(fundamnental niche)。因种间竞争,一种生物不可能利用其全部原始生态位,所占据的只是现实生态位(realized niche)。生态位概念应用于植物尚有一定困难。几乎一切植物都需要阳光进行光合作用,而且在陆地上倾向于占有环境的相同部分——土壤表面上下一定高度和深度。然而,植物种在自然环境水平地带和垂直地带中占据的区域以及在生长季节、开花季节等方面出现的差异,可与动物的生态位相比拟。
词义演变1910年,美国学者R.H.约翰逊第一次在生态学论述中使用生态位一词。1917年,J.格林内尔的《加州鸫的生态位关系》一文使该名词流传开来,但他当时所注意的是物种区系,所以侧重从生物分布的角度解释生态位概念,后人称之为空间生态位。1927年,C.埃尔顿著《动物生态学》一书,首次把生态位概念的重点转到生物群落上来。他认为:“一个动物的生态位是指它在生物环境中的地位,指它与食物和天敌的关系。”所以,埃尔顿强调的是功能生态位。 1957年,G.E.哈钦森建议用数学语言、用抽象空间来描绘生态位。例如,一个物种只能在一定的温度、湿度范围内生活,摄取食物的大小也常有一定限度,如果把温度、湿度和食物大小3个因子作为参数,这个物种的生态位就可以描绘在一个三维空间内;如果再添加其他生态因子,就得增加坐标轴,改三维空间为多维空间,所划定的多维体就可以看作生态位的抽象描绘,他称之为基本生态位。但在自然界中,因为各物种相互竞争,每一物种只能占据基本生态位的一部分,他称这部分为实际生态位。
后来R.H.惠特克等人建议:在生态位多维体的每一点上,还可累加一个表示物种反应的数量,如种群密度、资源利用情况等。于是,可以想象在多维体空间内弥漫着一片云雾,其各点的浓淡表示累加的数量,这样就进一步描绘了多维体内各点的情况。此外再增加一个时间轴,还可以把瞬时生态位转变为连续生态位,使不同时间内采用相同资源的两物种,在同一多维空间中各占不同的多维体;如果进一步把竞争的其他物种都纳入多维空间坐标系统,所得结果便相当于哈钦森的实际生态位。
计算公式生态位宽度 用多维空间描述生态位有助于概念的精确化。但实际工作中只能对少数几个,通常只是一两个生态因子作定量分析。生态位宽度是就一个生态因子轴而言的。这方面实验较多的是动物的竞争取食,以食物种类或体积大小为变量。设某物种在s种食物资源中取食,取每种食物的个体数分别为N1…Ni…Ns,则:

表示取食第i种食物的个体数在总数中的比例。R.莱文斯提出的计算生态位宽度B的公式是:

简单举例:设置装有不同食物的食槽s个,使一种动物取食,统计每槽取食的个体数。一种极端情况是,每槽个体数相等,表明该物种在所测范围内占有最宽的生态位,此时B值最大;另一种极端情况是,所有个体都在同一食槽取食,表明该物种在所测范围内占有最窄的生态位,此时B值最小。自然,这里测的是种群的综合效果。各个个体取食范围可能都很广(称泛化取食者),也可能都很窄(称特化取食者),但可因各特化取食者分别采取不同食物,所以它们的综合效果仍很广。但绝大多数情况介乎两者之间。
一般说来,当主要食物缺乏时,动物会扩大取食种类,食性趋向泛化,生态位加宽;当食物丰富时,取食种类又可能缩小,食性趋向特化,生态位变窄。
生态位重叠 生物群落中,多个物种取食相同食物的现象就是生态位重叠的一种表现,由此造成物种间的竞争,而食物缺乏时竞争加剧。R.莱文斯采用沃尔泰拉种群竞争公式中的竞争系数α表示重叠程度(见种群动态模型),用公式:
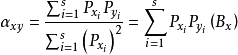
表示物种x对物种y的生态位重叠。以y的生态位宽度(By)代式中的(Bx),则得物种y对物种x的生态位重叠。公式表明:重叠程度取决于x和y取用同一食物的几率及生态位宽度。因为多数物种生态位宽度不等,所以重叠对两物种的影响也不相等,即可能:
生态位重叠与物种竞争密切相关,因此生态位概念是竞争排除原理的重要根据。
群落生境群落生境(其同义词为栖息地)只是生态位这个概念的一部分。生态位的含义远不止是“生活空间”(温度,空气湿度等环境因素的综合,它是生物生存的依据)的一个抽象概念,它描述了一个物种在其群落生境中的功能作用,而且它带有构成群落生境的自然因素所留下的烙印。它是一个物种为求生存而所需的广义“资源”。例如:蝙蝠需要在某地夜间捕食蚊子。这里面某地的自然因素(例如空气质量,其他关系到蝙蝠栖息地的因素),蝙蝠夜间运动的可行性,蚊子都是蝙蝠的生态位的一部分。一个物种只能占有一个生态位。
环境因素生态位的环境因素(温度,食物,地表湿度,生存环境等)的综合,构成概念生态位空间。这是一种n维超体积,但出于可视化的原因会将它简化为二维或三维龛位图进行显示。每种环境因素成为一个维度。在两个生态龛位中,考虑观察的维度越多,两个生态龛位的差别就越明显,越容易被区分开来。
分类基本生态位:是生态位空间的一部分,一个物种有在其中生存的可能。这个基本生态位是由物种的变异和适应能力决定的,而并非其地理因素。或者说基本生态位是实验室条件下的生态位,里面不存在捕食者和竞争。
现实生态位:是基本生态位的一部分,但考虑到生物因素和它们之间的相互作用。或者说是自然界中真实存在的生态位。
人们可以从特殊的性质或角度考虑,定义更多的生态位:
营养生态位: 根据营养情况划分的生态位。
最小环境: 对一个物种来说可持续生存的最小环境。
重合两个地区,虽然地理上被分隔,但却有着相似的非生物因素,在这两个地区生活的物种会占有相似的生态位。这会导致趋同演化的发生,即是两个物种虽然无亲缘关系,但却各自独立的发展出相似的身体构造去适应环境。南极的捕鱼能手,不会飞的鸟--企鹅和已灭绝的欧洲的大海雀一样占有相似的生态位。澳洲的袋鼹和欧洲的田鼠都有挖土铲的前肢,它们在泥土中挖掘通道,捕食细小的动物,它们占有相似的细小地下肉食性动物生态位。
生态平衡时,各个生物的生态位原则上不重合。若有重合,那么必然是不稳定的,它必然会通过物种间的竞争来削减生态位的重叠,直到平衡为止。竞争,比如需要相似生态位的入侵物种的进入,会导土著物种存在区域减少。如果存在区域太小,会导致一个物种的灭绝。这就是这就是竞争排除原则。
进化导致的是两个有亲缘关系的物种去占据不同的生态位,减少竞争的机会。一个很好的例子是拉帕戈斯群岛的达尔文雀的进化,(适应辐射)。
数量生态位的数量与生态系统的气候,地理和生物因素有关。相应的物种数会因这些因素的差异导致有很大的不同。极地,例如格陵兰的冰川,南极洲或是高原的生态位数就不及热带的原始森林或是珊瑚礁的多。
种群结构迄今,我们只处理了单一的局部繁殖种群或同类群:接下去,我们来考虑C—H—W平衡,突变、自然选择和随机遗传漂变以及它们在单一繁殖种群中的效应,可是,整个物种的种群很少会形成一个单一的大繁殖种群。取而代之,物种种群常以某种方式细分为许多分离的繁殖种群,如若不然,物种范围通常就太大,以至该范围内不同部分的个体,因为彼此为遥远的距离所隔离而没有机会交配。例如,马萨诸赛州的兔子就不与明尼苏达的兔子交配,因为垂直的物理距离分隔着它们。因此,种群结构或物种种群在空间上的分布方式,可能是重要的。
要讨论种群结构,必须引入几个附加名词。在对同类群或局部繁殖种群的比较上,我们将采用更普遍的孟德尔式种群的表达来表示以交配和家系纽带联系着的个体系统,或具有共同基因库的繁殖同伴。如此,一个孟德尔式种群便可看作是像一个同类群一样小或像整个物种一样大。多态变异表示一个同类群中的变异。而同地分布的这个词表示生活在同一故土,也表示同一同类群的成员或者由于彼此接近至少成为潜在偶配的那些个体。一个比较定量的定义是说,同地分布的石Qi体是指那些在一个个体及其所生后代的地点间的平均距离上被发现的有机体。这个定义是对个体的潜在配偶必然居住于其中的区域的一千粗略量度。异地分布意味着生活在另一故土,也衷东个体居住得如此遥远,以致它们不可能成为潜在的配偶。平行分布意味着居住在相邻区域,它表示地理上相邻的种群沿着接触带有可能交配。因此,多态变异是同类群内的变异,而不同的同类群遗传上常常彼此不同,故同类群间的变异常被称作多型变异。
种群结构的极端类型是连续型和不连续型。彻底的不连续分布叫做岛模型,该模型中的每个种群区别清晰、定义清楚且彼此孤立。诚然,它不仅可表示彼此由水分离着的岛屿种群,而且可表示占据一系列湖泊或池塘或坭坑或沼泽或被大草原环绕的小片林地或被树林环抱的小片草原上的种群。岛模型适合于对一个物种有利的生境区域及其类似区域被不利生境分隔开来的任何情况。连续分布则可在诸如北美洲北部的针叶林或东部的落叶林或西部大草原之类的大面积均匀生境中找到。虽然在表面上这类环境可表现为均匀的,而种群却不能是均匀的即使种群是均匀的,由于距离隔离,真正的随机交配也不可能达到2。
企业在生态学中,生态位是指在生态系统和群落中,一个物种与其他物种相关联的特定时间位置、空间位置和功能地位。生态位这一概念既表示生存空间的特性,也包括生活在其中的生物的特性,如能量来源、活动时间、行为以及种间关系等。生态位概念不仅指生存空间,它主要强调生物有机体本身在其群落中的机能作用和地位,特别是与其他物种的营养关系。因此,在自然界中,亲缘关系密切、生活需求与习性非常接近的物种,通常分布在不同的地理区域、或在同一地区的不同栖息地中、或者采用其他生活方式以避免竞争,如昼夜或季节活动上的区别、食性的区别等。生物在形成自身生态位的过程中遵循下述原则:趋适原则、竞争原则、开拓原则和平衡原则。趋适原则是指生物出于本能需要而寻求良好的生态位,这种趋适行为的结果导致生物所需资源的流动。竞争原则发生于不同生物之间对资源和环境因子的竞争。开拓原则是指生物不断开拓和占领一切可以利用的空余生态位。平衡原则是指作为一个开放的生物生态系统,总是向着尽力减小生态位势(竞争所导致的理想生态位与现实生态位之间的差距)的方向演替,因为一个生态位势过大的系统是一种不稳定的系统。上述的趋适和竞争原则解释了生物竞争行为的根源,开拓原则说明了生物可以主动的去改变自身状况,而平衡原则则说明每个生物在整个生态系统中都会找到一个最适宜生存的生态位,这就要求生物通过最恰当的竞争策略去寻找自身最恰当的生态位。

企业竞争企业界的情况与生物界十分类似,不同的企业也都有自己的“生态位”。许多规模和实力都很弱小的中小型企业也可以与那些可谓庞然大物的巨型企业在市场中共同生存和发展,其根本原因就在于它们拥有不同的生态位。
自然界中每个物种在进化过程中经过自然选择,形成特定的形态和功能,在生态空间中占有特定的生态位,形成生命系统的多样性。在自然生态系统中,只有生态位重叠的生命系统才会产生争夺生态位的竞争,竞争是争夺最适宜生存的生态区域。自然物种的生态位实际上是物种能获得和利用的生态资源空间,生态位越宽,物种的适应性越强,可利用的资源越多,物种的竞争力越强。
企业与自然生命系统不同,自然生命系统之间的竞争是在现有资源空间中争夺生存资源的竞争,自然生命不能创造其自身需要的资源空间,而企业一般来说在他的生命过程中没有特定的形态,他不但可以发展为经营任何行业与产品,并且可以创造出他生存所需的生存空间。因此,企业竞争生态位选择表现为在现有生态空间中的生态位选择和生态空间创新选择。
竞争策略1、错位经营策略
错位经营是指企业根据自身的资源组合和利用效率,选择区别于竞争
对手的发展重点进行生产和经营。企业在经营过程中,应该像生态位分离原理所昭示的那样,彼此错落有致,形成错位经营,默契地相互依存,相辅相成,从而避免价格战、产品战和广告战,寻求共同的发展。企业错位经营的主要内容有:(1)经营时间上的错位:企业在经营过程中应该像老鹰、猫头鹰实行白天和黑夜的时间错位。(2)经营空间上的错位:所有的商业、企业不一定都往繁华的市中心钻,应该像沼泽地中的药类那样,各自选择有利于自己发展的生存空间。(3)经营功能上的错位:包括经营规模、经营档次、经营业态、经营类别上的错位。所有的企业要根据当地购买力的不同情况,采取多种经营业态结合的合理布局,各自选择好自己的生态位,各就各位避开过度的残酷竞争。
2、生态位拓展策略
企业在发展的过程中,必须对自己的生态位进行控制和优化,以适应和改造生态环境。生态位控制包括两个方面的内容:拓展生态位和调整生态位。一个企业成功的发展必须善于拓展生态位和调整生态位,以改造和适应环境。只开拓不适应缺乏发展的稳度和柔度;只适应不开拓缺乏发展的速度和力度。企业在成长的过程中,对生态位的拓展速度要掌握好,避免过快,使企业发展失去连续性,在竞争中失败。
3、虚拟生态位策略
企业生态资源状况对企业生态位具有决定意义,随着人类社会由工业社会迈向信息社会,企业生态资源概念的外延、内涵及其企业对生态资源的占有方式正在发生深刻的变化。在信息社会,知识将作为占主导地位的资源和生产要素而存在,企业之间的竞争主要不是自然资源和其它有形的资本的竞争,而是拥有知识和技术多少的竞争、知识创新的竞争。信息社会竞争的加剧和环境变化的加快,使得企业无法单纯依靠自己生态资源来满足市场的需要。虚拟生态位和生态资源的共享为企业的发展拓展了生态空间,虚拟生态位的实质是发挥自身优势,对外部资源和力量进行有效整合,拓展自己的生态资源,达到降低成本、提高竞争力的目的。所以,企业要善于利用和建立虚拟生态位,以丰富自己的生态资源,改善生存、竞争环境,从而占据有利的生态位。巨大的全球网络、信息的光速流动使得企业生态资源的共享简单而便捷。
本词条内容贡献者为:
刘敦华 - 教授 - 宁夏大学
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国