

定义
有性生殖是通过生殖细胞结合的生殖方式。通常生物的生活周期中包括二倍体时期与单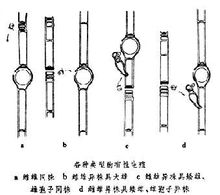 倍体时期的交替。二倍体细胞借减数分裂产生单倍体细胞(雌雄配子或卵细胞和精子);单倍体细胞通过受精(核融合)形成新的二倍体细胞。2这种有配子融合过程的有性生殖称为融合生殖。某些生物的配子可不经融合而单独发育为新个体,为单性生殖,是特殊的有性生殖,所以花药离体培养也属于有性生殖。
倍体时期的交替。二倍体细胞借减数分裂产生单倍体细胞(雌雄配子或卵细胞和精子);单倍体细胞通过受精(核融合)形成新的二倍体细胞。2这种有配子融合过程的有性生殖称为融合生殖。某些生物的配子可不经融合而单独发育为新个体,为单性生殖,是特殊的有性生殖,所以花药离体培养也属于有性生殖。
常见方式接合生殖单细胞生物有性生殖由个体直接进行,称接合生殖;2多细胞生物及单细胞生物的群体则由特化的单倍体细胞,即配子,进行融合生殖或单性生殖。 细菌的接合生殖是指两个菌体通过暂时形成的原生质桥单向的转移遗传信息。供体(雄体的部分染色体)可以转移到受体(雌体)的细胞中并导致基因重组。这是最原始的接合生殖。
细菌的接合生殖是指两个菌体通过暂时形成的原生质桥单向的转移遗传信息。供体(雄体的部分染色体)可以转移到受体(雌体)的细胞中并导致基因重组。这是最原始的接合生殖。
原生动物的接合生殖 多见于纤毛虫类,按接合的双方,即接合子的形态又可分为两类:
1)同配接合
接合子的形态相同。接合时双方暂时融合,小核在减数分裂后进行交换,相互受精后分开,如草履虫。
接合双方紧靠在一起,口部融合,然后大核消失小核分裂二次,成4个,其中3个退化,一个再经一次分裂成为一个动核和一个静核。此时接合个体互相交换动核,然后静核与换来的动核融合。接着虫体分开,每个个体的融合核分裂三次形成8个核,其中4个成大核,3个退化。此后大核不分裂,剩下的小核与虫体同时分裂两次而成为4个新的子体。
2)异配接合
见于缘毛目类纤毛虫。在进行接合生殖前,虫体先经一次不均等分裂,除小核外大核和虫体都分成大小两部分,成为大接合子和小接合子,前者固着,后者自由游泳。小接合子找到大接合子后即牢固附着在其上开始接合。在接合过程中,合子核只在大接合子中形成,小接合子为大接合子吸收。如钟虫。
配子生殖配子是由营养个体所产生的生殖细胞,需两两配合后才能继续其生活史,如在一定时间内找不到适当的配子便死亡。按配子的大小,形状和性表现可分为三种类型:
1)同配生殖:配子的形态和机能完全相同,没有性的区分。例如衣藻属中的大多数种类。
2)异配生殖:有两种类型:①生理的异配生殖,参加结合的配子形态上并无区别,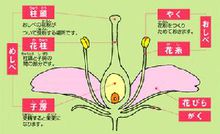 但交配型不同,在相同交配型的配子间不发生结合,只有不同交配型的配子才能结合,且具有种的特异性。如衣藻属中的少数种类。这是异配生殖中最原始的类型。②形态的异配生殖,参加结合的配子形状相同,但大小和性表现不同。大的不大活泼为雌配子,小的活泼,为雄配子,这说明已开始了性在形态上的分化。
但交配型不同,在相同交配型的配子间不发生结合,只有不同交配型的配子才能结合,且具有种的特异性。如衣藻属中的少数种类。这是异配生殖中最原始的类型。②形态的异配生殖,参加结合的配子形状相同,但大小和性表现不同。大的不大活泼为雌配子,小的活泼,为雄配子,这说明已开始了性在形态上的分化。
3)卵配生殖:相结合的雌雄配子高度特化,其大小、形态和性表现都明显不同,成为卵和精子。卵和精子经过受精,融合为受精卵。卵配生殖是分化显著的异配生殖。
配子生殖的进化趋势是由同配到异配,最后发展为卵配生殖。在原生动物和单细胞植物中,所有个体或营养细胞都可能直接转变为配子或产生配子,而在高等动物中,生殖细胞是由特殊的性腺产生的。
起源有性生殖是如何起源的(即为什么有性)?这样一个看似简单的问题不仅令达尔文困惑不已,150年过去了, 人们都还未找到一个普遍认可的理论, 这也被称为是进化生物学问题之皇后“ queen of problems in evolutionary biology” (Bell, 1982)。法国著名的遗传学家、诺贝尔生理学或医学奖(1965年)获奖者Jacob (1998)在其专著《鼠、蝇、人与遗传学》一书中感叹道: “ 在通过有性方式来进行生殖的生物中, 一切都被安排得要使同一物种的全部个体(除了真正的孪生个体)都互有差异。这就犹如整个地球上起作用的遗传系统已被调节得要永远产生差异。因此就有了这个悖论: 一方面, 所有显得十分不同的东西, 归根结底却是很相似; 另一方面, 所有显得十分相似的东西, 事实上却是相当不同” 。《为什么要相信达尔文》一书的作者Coyne (2009)感叹道: “ 为什么会演化出性别来?这其实是演化论最大的谜团之一” 。
有性生殖发生的直接证据,最早见于澳大利亚中部的苦泉燧石中,在这里发 现了植物减数分裂产生的四分孢子的化石。岩石的年龄约为10亿年,估计有性生殖实际出现还要早些,约在真核生物产生后不久。从动、植物生殖细胞的形成过程中均有复杂的减数分裂来看,有性生殖应起源于动、植物分化前,但这还只是一种推测。
现了植物减数分裂产生的四分孢子的化石。岩石的年龄约为10亿年,估计有性生殖实际出现还要早些,约在真核生物产生后不久。从动、植物生殖细胞的形成过程中均有复杂的减数分裂来看,有性生殖应起源于动、植物分化前,但这还只是一种推测。
而关于性是如何起源的更是知之甚少,从细胞遗传学的角度,人们提出了共生起源学说、细胞分裂误记说和DNA纠错说等(德迪夫1999,洛耶 2004,威廉斯2008)。谢平(2013)认为,在很多动植物中,“性”往往与休眠现象联系在一起,产生后壁孢子、种子或休眠卵,因此提出了有性生殖的休眠起源说(accident dormancy hypothesis),但分子机制有待进一步揭示3。从分子遗传学的角度来说, “性”一点都不神秘, 就是将两个个体的基因组拼在一起而已, 藉此种族多样的遗传信息分散到了个体之中;而从生态的角度来看, “性”的原始动机就是与休眠事件的偶联4。
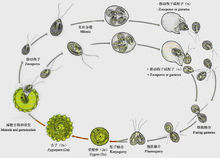
意义有性生殖中基因组合的广泛变异能增加子代适应自然选择的能力。有性生殖产生的后代中随机组合的基因对物种可能有利,也可能不利,但至少会增加少数个体在难以预料和不断变化的环境中存活的机会,从而对物种有利。
有性生殖还能够促进有利突变在种群中的传播。如果一个物种有两个个体在不同的位点上发生了有利突变,在无性生殖的种群内,这两个突变体必将竞争,直到一个消灭为止,无法同时保留这两个有利的突变。但在有性生殖的种群内,通过交配与重组,可以使这两个有利的突变同时进入同一个体的基因组中,并且同时在种群中传播。
此外进行有性生殖的物种其生活周期中都有二倍体的阶段。二倍体的物种每 一基因都有两份,有一份在机能上处于备用状态。如果这个备用的基因发生突变,成为有新的功能的基因,但此时新功能还是潜在的。通过自发的重复和有性生殖中的遗传重组,这个新基因可与原有基因先后排列,这样便产生一个新的基因。二倍体物种可以用这样的方法使其基因组不断丰富。
一基因都有两份,有一份在机能上处于备用状态。如果这个备用的基因发生突变,成为有新的功能的基因,但此时新功能还是潜在的。通过自发的重复和有性生殖中的遗传重组,这个新基因可与原有基因先后排列,这样便产生一个新的基因。二倍体物种可以用这样的方法使其基因组不断丰富。
由于上述原因,有性生殖加速了进化的进程。在地球上生物进化的30余亿年中,前20余亿年生命停留在无性生殖阶段,进化缓慢,后10亿年左右进化速度明显加快。除了地球环境的变化(例如含氧大气的出现等)外,有性生殖的发生与发展也是一个主要的原因。现存150余万种生物中,从细菌到高等动植物,能进行有性生殖的种类占98%以上,就说明了这一点。
其它方式动物的有性生殖中除精卵结合外,尚有一些其它形式,如:
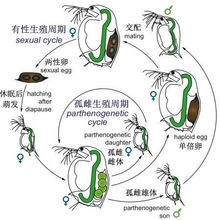
孤雌生殖 孤雌生殖(parthenogenesis):有些动物的生殖不需雄体参加,只是雌体产生卵。此卵不与精子结合,即可发育成新的个体,实际上这是单性生殖。其卵发育成的子代一般均为雌性个体。少数动物至今只有雌性个体,尚未发现雄性个体的存在,一生中只进行孤雌生殖,如某些轮虫。有些动物的生殖是两性生殖与孤雌生殖并存,如蜜蜂,其卵不受精即发育成雄蜂;卵受精则发育成雌性的工蜂和蜂王。有的动物在一生中有一段时期进行两性生殖,另一时期进行孤雌生殖。如蚜虫,在其生活史中,大多数时间进行孤雌生殖,只到秋末时才进行两性生殖,产生的卵与精子结合后,休眠越冬,次年再发育成雌性蚜虫。
孤雌生殖(parthenogenesis):有些动物的生殖不需雄体参加,只是雌体产生卵。此卵不与精子结合,即可发育成新的个体,实际上这是单性生殖。其卵发育成的子代一般均为雌性个体。少数动物至今只有雌性个体,尚未发现雄性个体的存在,一生中只进行孤雌生殖,如某些轮虫。有些动物的生殖是两性生殖与孤雌生殖并存,如蜜蜂,其卵不受精即发育成雄蜂;卵受精则发育成雌性的工蜂和蜂王。有的动物在一生中有一段时期进行两性生殖,另一时期进行孤雌生殖。如蚜虫,在其生活史中,大多数时间进行孤雌生殖,只到秋末时才进行两性生殖,产生的卵与精子结合后,休眠越冬,次年再发育成雌性蚜虫。
幼体生殖幼体生殖(paedogenesis):动物的幼体生长发育成熟后,即形成成体,其生殖器官才能产生精子或卵,进行有性生殖。但某些动物的幼体虽未发育为成体,即未达性成熟,在幼体阶段就可进行生殖,繁殖后代,称为幼体生殖。幼体生殖产生的不是卵而都是幼体,因此这可以认为是胎生的一种形式。幼体生殖的母体在进行生殖时不需雄体参加,即卵不与精子结合,因此幼体生殖也可看成是孤雌生殖的一种类型。如瘿蚊(Oligarcesparadoxus)除夏季进行两性生殖外,其余季节都进行幼体生殖。
多胚生殖多胚生殖(polyembryony):一个卵可产生二个或更多的胚胎,发育成多个个体的生殖方法,称为多胚生殖。此种生殖为卵核分裂一次,以后就形成两个胚胎,但大多数种类的卵核都经多次分裂,形成许多子核。多者可产生1600~1800个。如广腹细蜂(Platygasterrernalis)一个卵能产生15~20个胚胎;小蜂(Litomastixtruncatellus)可产生2000个胚胎。
对比无性生殖与有性生殖的比较
·无性生殖
无性生殖——显微镜下的结构不经过生殖细胞的结合,由母体直接产生出新个体的生殖方式叫无性生殖。无性生殖的方式:
1、分裂生殖,如变形虫;2、出芽生殖,如水螅;3、孢子生殖,如根霉;4、营养生殖,如草莓。扦插、嫁接都属于营养生殖。如:“无心插柳柳成荫”。克隆的原意是“离体的小树枝发育成一个植物体。”即无性繁殖的意思。高等植物的无性繁殖比较普遍,而高等动物的无性繁殖则比较困难,1997年英国科学家用克隆技术培育出了“多莉羊”,这标志着生物新世纪的到来。
有性生殖
由亲本产生有性生殖细胞(也叫配子),经过两性生殖细胞(如卵细胞和精子)的结合,成为合子(如受精卵),再由合子发育成为新个体的生殖方式,叫做有性生殖。有性生殖有三种:同配生殖、异配生殖和卵式生殖。而配子生殖中最高级的生殖方式是——卵式生殖。
团藻是生活在淡水中的一种能够游动的藻类植物。团藻的直径只有1-2毫米。一个团藻由几百个到几万个细胞构成,这些细胞都排在团藻的表面,其中只有少数的细胞具有生殖的功能。在进行有性生殖时,有的细胞长得很大,失去鞭毛,不能游动,这种大的配子叫卵细胞;有的细胞能够产生大量的小细胞,小细胞生有两根鞭毛,能够游动,这种小配子叫做精子。两者大小悬殊。
精子成熟时,游到卵细胞近旁,与卵细胞结合成合子(这种合子也叫做受精卵),合子外被厚壁,这种合子可以发育成新的个体——团藻。这种卵细胞与精子结合的生殖方式叫卵式生殖。
在单细胞中普遍的生殖方式是分裂生殖;生物界中普遍的生殖方式是有性生殖;高等植物和多数动物所普遍具有的一种有性生殖方式是卵式生殖。
·对比
1、无性生殖和有性生殖的根本区别是:前者不经过两性生殖细胞的结合,由母体直接产生新个体;后者要经过两性生殖细胞的结合,成为合子,由合子发育成新个体。
2、有性生殖与无性生殖相比具有更大的生活力和变异性。从进化的观点看生物的生殖方式是由无性生殖向有性生殖的过渡。
评价有性生殖通过生殖细胞的生殖。通常生物的生活周期中包括二倍体时期与单倍体时期的交替。二倍体细胞借减数分裂产生单倍体细胞(雌雄配子或卵和精子)。细菌的接合生殖是两个菌体暂时形成原生质桥由供体向受体单向的转移遗传信息,并导致基因重组。原生动物接合时双方(称接合子)暂时融合,小核在减数分裂后进行交换,相互受精后分开。
有性生殖中基因组合的广泛变异能增加子代适应自然选择的能力。有性生殖能促进有利突变在种群中的传播。有性生殖的物种其生活周期中都有二倍体的阶段。二倍体的物种每一基因都有两份,有一份在机能上处于备用状态。由于上述原因,有性生殖加速了进化的进程。现存150余万种生物中,从细菌到高等动植物,能进行有性生殖的种类占93%以上,似乎说明了这一点5。
很多人认为两性生殖必定优于单性生殖,只是争议为何两性生殖优于单性生殖。为此,人们已经提出的关于两性生殖优于单性生殖的假说多达二十多种,Kondrashov(1993)对这些假说进行了归类,如表1所示。些假说带有浓厚的遗传学背景,普遍相信有性生殖能够有效地积累有益突变与去掉有害突变。
表1 关于两性生殖优于单性生殖假说的分类
Table 1 A classification of hypotheses on the advantage of amphimixis over apomixis
I.直接有益假说Immediate benefit hypotheses
1. 增加后代的适合度 因为二倍体后代来自两个而不是一个亲本
因为双链DNA损伤在减数分裂的染色体结合中能够得到修复
2. 降低有害突变速率 因为许多突变源自缺失(deletion),而这在染色体结合时能作为缺口(gap)而被
识别,能通过基因转换偏性(biased gene conversion)来填补
因为许多有害突变源自插入(insertion),它在染色体结合过程中作为环(loop)
能被识别和被切除
因为许多表突变(epimutation)源自去甲基化(demethylation),这在染色体结合
过程能被识别和恢复
3. 增加选择效率 因为在雄性或雄配子的选择可能代价较小
**II.****变异和选择假说(局域种群)**Variation and selection hypotheses (local population)
1. 在随机性环境假说中 在许多基因座,有益突变能更快速的同步累积
在有害突变存在时,能更快速地积累有益突变
当多态(polymorphisms)保持在其它基因座时,能更快速地积累有益突变
2. 在确定性环境假说中 在不可逆变化的环境中,具有更快的进化速率
在宽幅波动的环境中,种群具有较快的变化速率
在频繁波动的环境中,种群的变化速率较低
3.在随机式突变假说中 非有害突变基因型随机丧失的恢复
4. 在确定式突变假说中 提高了针对协同作用有害突变(synergistically interacting deleterious
mutations)的选择效率
**III.****变异和选择假说(空间结构种群)**Variation and selection hypotheses (spatially structured population)
1. 在随机的联系中 由于遗传差异,它防止了寄生虫从母体向子代的传播
它增加了一类亲缘动(植)物共有最佳基因型的机会
它降低了亲缘动(植)物间的竞争
2. 在确定的联系中 它组合了在不同亚种群形成的不同的有益等位基因
(引自Kondrashov 1993)(cited from Kondrashov 1993)
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国