

甲壳亚门是节肢动物门中的一个亚门,由于亚门下物种用鳃呼吸,过去曾与三叶虫纲一起列为有鳃亚门(Branchiata),后来又被列为有颚亚门(Mandibulata),后有更多学者主张应独立成甲壳亚门(Crustacea)。甲壳类动物约有31000多种,包括我们熟悉的水蚤、剑水蚤、丰年虫、对虾、螯虾、龙虾、蟹等。
简介甲壳亚门在形态结构上表现出多样性,例如体形大小、体节数目、体躯划分及附肢的分化都有较大的差异。在生活方式上绝大多数为海洋生,为海洋浮游动物的重要成员,常被誉为海洋中的昆虫,少数侵入淡水。是构成水生食物链中的重要环节。
也有极少数进入陆地生活,但不能脱离潮湿环境。
甲壳类动物都具有两对触角,这一点区别于其它节肢动物(有螯肢亚门无触角,三叶虫亚门及单肢亚门仅有一对触角,相当于甲壳类的小触角)。甲壳类动物的附肢基本上都是双肢型,但在不同的体区有不同的形态与机能的分化,其中至少有三对口后的附肢作为颚。
甲壳类动物生活史中由卵孵出后为无节幼虫(nauplius),仅有三个体节,三对附肢,相当于成体的前三对附肢,即两对触角及一对大颚。经多次蜕皮不断地在肛门前部增加体节数,原始的种类体节可多达数十节,高等的种类体节数减少并固定,体区划分清楚。
此外有些种类头部和胸部可能也会融合到一起,形成头胸部。
所有的甲壳亚门的干细胞分裂时的特征相同。
包含分类甲壳亚门分为6个纲(Martin&Davis,2001)。为叙述方便常将小型甲壳动物(前5个纲)统称切甲类(Entomostracans),将大型的(软甲纲)称为软甲类(Malacostracans)。但这种命名不具分类意义。
六纲为:
鳃足纲
桨足纲
头虾纲
颚足纲
介形纲
软甲纲
外部形态甲壳动物的形态有着较多的变化,随不同的种类其体节、分区都有很大的差异。
低等的甲壳类一般体型小,只有几mm到2~3cm,体节数多且不固定,但分节明显,各节形态相似,体表外骨骼薄而透明。例如丰年虫(Chirocephalus),身体分为头部与躯干部,没有背甲。又如鲎虫(Apus),体节数可多达50多节,胸、腹部分界不明显,有背甲(carapace),其头部体节与胸部体节愈合成头胸部(cephalothorax)。还有的种类体表完全被石灰质骨板包围,并营固着生活,例如藤壶(Balanus)。低等甲壳类生物形态的多样性在细分的纲中会有较为详细的介绍。1
高等的甲壳类动物,即软甲纲动物身体的节数固定,分区明显,一般是由20~21节组成。例如虾的头部5节,胸部8节,腹部6节及1尾节。部分胸部体节与头部合并,形成头胸部,之间分界消失。头胸部背面具有一发达的背甲,也称头胸甲,盖在身体的背部,或向两侧延伸也盖住体侧的部分附肢及鳃,因此也称为鳃盖,其下的空腔称鳃腔。头胸甲是由头及胸部的体褶共同分泌的外骨骼所形成。
甲壳类动物有发达的附肢,除了触角与尾节之外,每个体节都有一对双肢型附肢。随着不同种及不同的着生部位,附肢有多种形态及功能的分化。原始的种类附肢数目随体节数也较多,但分化很少,多呈叶片状(foliaceous),它相似于环节动物的疣足,与体躯无关节而是直接相连,可分为内叶(内肢)(endopodite)与外叶(外肢)(exopodite),基部的突起形成了副叶(epipodite),具鳃的功能。丰年虫、鲎虫都是这种原始的叶片状附肢。
高等的甲壳类动物附肢随体节数减少而减少,均与躯干部以关节相连,附肢本身亦分节。与躯干相关节的部分称原肢节(protopodite),由原肢节再生出内肢与外肢,虾、蟹腹部的游泳足便是这样。多数附肢、特别是胸部附肢原肢节可分为两节,与体躯相关节的称为底节(coxopodite),其下一节为基节(basipodite)。由基节再分出内肢与外肢。内肢可再分成5节,由基部到端部依次为座节(ischiopodite)、长节(meropodite)、腕节(carpopodite),掌节(propodite)及指节(dactylopodite)。原肢节上可长出旁枝,称副肢。在原肢节的内外边缘还可长出突起,称外小叶(exite)及内小叶(endite),一些种类外小叶变成了鳃,靠近口部附肢的内小叶变成了颚。如果外肢节减小或退化,则变成单肢型附肢。
以沼虾(Macrobrachium)为例,共有19对附肢,它们的分布是:头部5对,其中第一对为小触角(antenn-ules),单肢型,同源于其他亚门的触角,第二对为大触角(antennae),均具感觉功能。第三对为大颚(mandibles),其原肢节大而坚硬,内面具齿,用以咀嚼。后二对为第一及第二小颚(maxillae),呈叶片状,用以把持食物,第二小颚的外肢扩大形成颚基叶(gnathobasic lobes),用以激动水在鳃腔中的流动。后三对附肢位于口后。口的前、后有体壁形成的突起,构成上唇(labrum)及下唇(labium)。胸部8对附肢,前三对形成颚足(maxillipeds),双肢型,具副肢及鳃,用以呼吸、感觉及把持食物。胸部的后5对为步行足(pereiopoda),外肢退化或消失,原肢2节,内肢5节,末端具螯或爪,具步行、取食、呼吸及防卫的功能。腹部6对附肢,原始的双肢型,原肢节及内肢节均不分节,用以游泳,称腹足(pleopoda)。雌性的可用以携卵,雄性的第1~2对附肢用以输送精子。第6对附肢与尾节合并成尾扇,具舵的功能。
体壁与色素体壁结构与其它节肢动物相似,只是某些甲壳类如虾、蟹的外骨骼有相当程度的钙化,主要是因为上表皮及外表皮均有相当多钙盐的沉积。另外在表皮细胞的下面有皮下腺(tegumentalgland)和色素细胞(ehromatophores)。
皮下腺是一群具分泌能力的腺细胞,并有长管穿过外骨骼开口在上表皮的表面,它的功能还不十分清楚。色素细胞是位于表皮细胞下面的结缔组织中的一些呈放射状分支的细胞,放射状的突起不能伸缩,但细胞中含有大量的色素颗粒,呈白、红、黄、蓝、褐、黑等色。根据生理状况的不同,颗粒或分散在整个细胞,或集中在细胞中心。其中红、黄、蓝色是类胡萝卜素,来自于食物,在生活状态时,这些色素常与蛋白质呈结合状态而表现出不同的颜色。
当虾、蟹加热煮熟时,体表出现美丽的红色,这是因为加热使蛋白色沉淀出来而留下红色化合物——虾青素(astaxanthin)的缘故。事实上,一个单个的色素细胞可以包括一种到几种不同颜色的色素,任何一种色素都可独立移动。多色素的色素细胞仅出现于虾中。
体色的改变有两种形式:一种是形态变色(morphologicalcolorchange),它涉及色素细胞中色素的丢失或形成,或是由于动物长期在一固定的环境及光照条件下细胞数目的改变而形成;另一种形式是生理变色(physiological color change)它是指对环境的迅速的颜色适应,来自于色素细胞中色素颗粒的分散与集中。
最普通的生理变色是体色深浅的改变,这在许多蟹中可以发现。而虾类常有更广泛的颜色变化,例如一种小长臂虾(Palaemonetes),色素细胞中含有红、黄、蓝三种色素颗粒,通过其中任一色素颗粒的独立移动使之可以适应任何颜色的背景。许多虾类都具有这种能力。色素颗粒在色素细胞中的移动受眼柄中分泌的激素(hormones)控制。例如许多虾的色素细胞含有红、黄、蓝和白色素,移走眼柄,红、黄色素扩散而使体色变暗。如注射眼柄的激素提取物,则白色素扩散而使体色迅速变浅。
因此,对每种色素可能都存在着一对对抗性的促色素细胞素(chromatophorotrophins),其中一个处于分散状态,另一个处于集中状态。体色深浅的改变,取决于一对对抗性促色素细胞素中的哪一个处于扩散状态以及哪一个处于集中状态。
运动与取食甲壳类动物的运动方式与生活方式密切相关,小型的甲壳类动物保留了祖先的水底表面悬浮取食的习性,因此它们以某些附肢的打动以推动身体前进。例如水蚤(Daphnia)、剑水蚤(Cyclops)等以第二对触角像桨一样在水中划动,推动身体呈跳跃式的前进。在水中游泳生活的种类,其游泳肢具有刚毛繸,以增加抗水表面。
大型游泳生活的甲壳类动物腹部具有发达的肌肉,靠肌肉的收缩推动身体前进,例如各种虾。
爬行生活的蟹类,虽然某些种还保留了游泳的能力,但更多的附肢已变得适合于爬行及挖掘。
甲壳类动物的食物类型及取食习性也表现出多样性,有植食的、肉食的、腐食的以及寄生生活的,取食方式及相应的附肢因种而有很大不同。许多小型的甲壳类动物都是过滤取食,它们以浮游生物或沉渣为食,这种取食方式可能与运动相关。其头部及躯干前端的附肢布满细微的刚毛,不仅用以激动水流,同时也作为过滤器以收集水中的食物颗粒。刚毛间空间的大小决定了食物颗粒的大小。刚毛收集的食物颗粒经附肢腹面的食物沟送入口。
大型的甲壳类动物多为捕食性取食,前端的步足适合于捕获及撕裂食物,大小颚及颚足用以咬切及把持食物。大小颚及上、下唇组成口器。十足目的第三对颚足排列在口外组成门状的保护器。消化道呈直管状,口位于前端腹面,前肠包括口、食道、膨大的贲门胃(cardiac stomach)及幽门胃(pyloric stomach)。在贲门胃及幽门胃的胃壁上有几丁质硬化形成的嵴及独立的齿,用以研磨食物,并有肌肉控制胃壁及齿的运动。
取食微小食物的种类嵴与齿不发达。幽门胃的胃壁多褶皱而使胃腔减小,褶皱上生有大量的刚毛而形成过滤系统,使小的食物颗粒进入消化盲囊,大的颗粒进入中肠。幽门胃后为中肠,中肠的前端有一对或数对消化盲囊。在十足目一对消化盲囊变成实体的消化腺,称为肝胰腺(hepatopancreas),它几乎充满了头胸部,它能产生消化酶并进行食物的消化,也是糖元、脂肪及钙的吸收与贮存的场所。中肠也是消化及吸收的场所。中肠后经后肠以肛门开口在尾节腹面。
呼吸小型甲壳类动物没有专门的呼吸器官,而是用体表进行气体交换。某些种虽有附肢特化成鳃,但仍以体表呼吸为主,例如鳃足类。
软甲纲都以鳃作为呼吸器官。典型的鳃与胸部体节的侧壁及附肢相关,鳃的数目、结构、位置因种而不同。原始的胸部每个体节应有4对鳃,其中一对附着在体侧壁上,称侧鳃(pleurobranch),2对固着在附肢底节与体壁相连的关节膜上,称关节鳃(arthrobranch),一对附着在附肢基节上,称足鳃(podobranch)。但现存种类实际上都少于32对鳃,例如深对虾(Benthesicymus)只有24对,螯虾只有17对,海产蟹类一般只有9对,最少的豆蟹(Pinnotheres)只有3对。所有的鳃均位于鳃室中,并被头胸甲延伸形成的鳃盖(branchiostegite)所遮盖。生活时靠附肢、特别是第2颚足的颚基叶的打动造成水流。水可由鳃盖的腹缘及后缘流入鳃室,或由附肢基部及鳃盖后缘流入。
在蟹类,仅由螯肢的基部流入。水都由头胸甲前端两侧流出。水在鳃腔中的循环时,使鳃得以进行气体交换。蟹类三对颚足副肢及部分步足的基节上具有大量的刚毛,用以清洁鳃及过滤水中的杂质,以防鳃被沉渣污染。
鳃的结构可分为三类:支鳃(dendrobranch),由鳃轴向两侧伸出侧支,或侧支再分支,例如对虾类;毛鳃(trichobranch),围绕鳃轴具毛状或丝状排列,例如大多数的长尾虾及螯虾等;叶鳃(phyllobranch),沿鳃轴向两侧伸出叶片状鳃页,例如长臂虾(palaemon)、寄居蟹类及蟹类,鳃轴中有入鳃血窦及出鳃血窦,流经鳃丝或鳃叶时进行气体的交换。
陆生及半陆生的种类仍然保留了较大的气体交换的表面积,例如海岸边生活的招潮蟹(Uca);它携带水分在鳃腔中,并在第3~4对足上有呼吸孔,以允许气体进入,气体的交换仍在鳃腔中进行。真正陆生的种类如椰子蟹(Birgus)、鼠妇(Porcellio)等,其鳃腔壁变成囊室而形成肺。
循环切甲类一般没有循环的结构,血液在血腔中通过身体的运动而循环。
软甲类都有循环系统,也属开放式,包括心脏、血管及血窦。心室的形态因种而异,或呈囊状、管状,或呈实体状,均位于胸部背面的围心窦中。心室上有心孔,血液由心孔进入心室。低等的种类,血液由心室出来,经短而不分支的动脉进入组织,再经静脉血窦进入鳃,行气体交换后再回到围心窦,最后回到心室。十足目的心室呈实体状,有三对心孔。
虾的心孔在心室的前侧角、后侧角各一对,背面一对。由心室向前发出五条动脉;一条眼动脉(ophthalmic artery);一对触角动脉(antennary arteries)及一对肝动脉(hepatic arteries)。由心脏向后发出一条腹动脉(abdominalartery),沿背中线后行。此外,还有一条胸直动脉(sternal artery),由心脏下面发出,沿消化道侧面直达腹面穿过神经索分成两支,称为神经下动脉(subneural artery),一支向前,一支向后。这些血管再分支,进入组织,然后流入胸窦,进入鳃后行气体交换,最后再流回围心窦,经心孔再流回心室。
甲壳类的血浆中溶解有血青蛋白,故血液多呈淡蓝色。血液中含有透明的及大的颗粒状的变形细胞,其数目因种、年龄及生理状态而不同,但一般在数百—数千/cm3血液中。血细胞不仅具吞噬能力,也参与血液的凝结。在刺激条件下,如断肢时某些变形细胞可以分解、破裂,并释放出某些物质将血浆中的纤维蛋白原(fibrinogen)转变成纤维蛋白(fibrin),结果出现凝结的血浆岛并被沉淀,形成血块。一些小型的甲壳类在血液及组织液中含有少量血红蛋白,它与氧的亲和力较大,适合于在低氧条件下呼吸。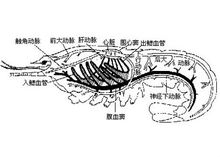
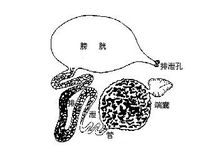
排泄甲壳类的排泄器官是一对触角腺(antennal glands),或是一对小颚腺(maxillary glands),因位于第二触角或小颚基部而得名。幼虫期二种腺体常同时存在,成虫期往往只有其中的一种。也有两种同时存在的种类,如某些介形类。其结构相似于有螯类的基节腺,也是由残留的按节排列的后肾形成。由基部的囊及排泄管组成。
在软甲类,触角腺或小颚腺有了高度的发展,端囊包括一个小囊(它从周围的血液中收集含废物的液体成分)及一个大的腺体部分,它是囊壁褶皱形成的海绵状组织,是重要的重吸收部位。由腺体部再通出排泄管,其后端膨大形成了膀胱,最后经一短管在第二触角的基部开口到体外。
甲壳类的代谢产物主要是氨及少量的尿酸。虽然触角腺称为排泄器官,但大量的含氮废物是通过鳃或体壁的扩散作用而排出体外。另外,在鳃轴处及足的基部有一些肾细胞(nephrocytes),这些细胞可以摄取及贮存尿酸形成颗粒,成为色素细胞的一部分,或沉淀在体壁中,随蜕皮时排出。而触角腺的主要机能在于维持及调节体内离子浓度及液体压力的平衡。当体内血液中水分含量增高时,触角腺的过滤作用也增强,并形成较多的尿而排出体外。例如一种蟹Carcinus,如果生活在海水中(含盐量34‰),每日产生相当于3.6%体重的尿,如果生活在咸水中(14‰),则每日产生相当于1/3体重的尿。这说明触角膜的机能在于维持稳定的体积常数,而不管外界环境中的盐度含量。
大多数十足目所产生的尿与血液是等渗的,而且是能调节盐度的,因为它们的鳃可以由水流中摄取离子以补偿由尿中失去的离子。另外,排泄管也具有重吸收作用,以产生低渗的尿排出体外。
神经与感官甲壳类的神经系统也是链状神经。原始的种类脑后的神经索呈双链状,例如某些鳃足类,即在每一个体节各有一对神经节,彼此分离,之间有成双的横神经相连,前后神经节各自成链,因此形成双链。
但大多数甲壳类不存在这种原始的结构,而是双链愈合成单链。如各种虾、蟹类胸腹部神经节常与食道下神经节合并成团块状,脑神经节也更集中。由脑发出神经到触角、眼,围咽神经环等。腹神经索中的每个神经节分出三对侧神经,其中前两对是运动与感觉神经,支配附肢与体节,最后一对是运动神经,支配体壁的肌肉。许多甲壳类的神经索中也有巨大神经纤维,能快速传导冲动产生反应。其纤维的直径可达200μm。一般在神经索的背中、背侧各有两个巨大纤维,前者是由脑神经细胞发出,其纤维达神经索的全长,后者是由神经节的细胞发出,所以刺激任何一种巨大纤维都可引起身体的快速运动。虾正是以这种机制完成快速跑动、取食、繁殖、自切等各种反射活动的。
甲壳类具两种类型的眼,即中眼(median eyes)与复眼(compound eyes)。中眼一个,是无节幼虫期所普遍有的,成体后或者存在,或者消失。在低等的种类成为成虫期仅有的眼,如桡足类。它由3~4个具色素杯的小眼组成,小眼由少量的视觉细胞组成,一般没有晶体状结构。中眼的机能可能仅在于测知光源,使动物趋向于水的表层或低层。大多数成体具一对复眼,分别位于头的两侧。多数种类眼位于眼柄(peduncle)上,眼柄由2~3节组成,一般可以活动。眼的表面角膜有很大的凸度,其弧面可达180°,再加上眼柄的运动,因此具有很大的视觉范围。复眼中的小眼数少的仅有20多个,多的可达15000个,如螯龙虾(Homarus)。
大多数甲壳类的复眼对强、弱光均有色素屏幕效应,也就是说,生活在光照条件下的种类(陆地、浅水)一般具并列眼,而一些洞穴及深水生活的种类由于色素屏幕不发达而具重叠眼。另外,从眼的结构研究表明,甲壳类的复眼能够识别物体的形状、大小及颜色变化。例如寄居的蟹类可以区别不同颜色的螺壳,一些虾可以在红、黄、褐色的背景中生存,而不适合深色环境。
十足目常有一对平衡囊,位于触角的基部,或足的基部或尾节,它由外胚层凹陷形成,内有平衡石,外有开口。平衡石或由外界摄入,或由细胞分泌。平衡囊也随蜕皮而脱落,蜕皮后再形成。囊的底部内壁有感觉毛,或与平衡石直接接触,或通过囊内的液体而感受刺激。平衡囊是其重力感受器,并与运动相关,以调节身体平衡。除了以上结构,甲壳动物外骨骼的表面分布有各种感觉毛,有多种机能。最常见的是分布于附肢上的触觉感受器,用以测知水流及其方向,它与外界环境直接接触。还有存在于触角及围绕口部附肢上的化学感受器,特别多地分布在小触角的外鞭上,由触角上的双极神经细胞所支配。分布于触角的还有温度感受器。在腹部肌肉两侧,还有一些由肌肉细胞改变形成的本体感受器(proprioceptors),它们的机能还不十分清楚。
生殖与发育大多数的甲壳动物是雌雄异体,但也有一些种类为雌雄同体,例如蔓足类、寄生的等足类等。还有一些种类为孤雌生殖,例如鳃足类、介形类等。
甲壳动物的生殖腺(精巢、卵巢)通常是一对长形器官,位于胸部或腹部背面,或同时存在于胸腹部。精巢一对,背部靠近或愈合成一整块。一对输精管。有的种类以精荚进行传递精子,这时其输精管的后端部分变成腺状,以便于精荚的形成。在十足类输精管末端形成肌肉质的射精管(ejaculatoryduct),最后以生殖孔开口在最后一对胸部附肢的基部,或最后胸节的腹板上。
十足目雄性的第1~2对腹足特化,用以传导精子。大多数甲壳类的精子是无尾的,呈球形或星形等非典型精子形态。卵巢的结构与位置相似于精巢。一对输卵管。十足目的输卵管末端联合,还形成受精囊及阴道,最后以雌性生殖孔开口在附肢基部。受精囊开口到外界,并在最后1~2对胸节腹板突起形成一盘状。
在十足目中出现某些种的性二型现象。例如招潮蟹,其雄性的一个附肢的爪特别发达,而雌性具二爪且相等大小。有的种雌雄个体大小相异。交配前也常有性引诱(sexual attraction)及追逐行为(courtship)。引诱行为或是视觉的,或是听觉的。例如,招潮蟹在交配之前雄性在雌性面前不断的摇动,其摇动的方式在同一种是固定的。另外,雄性的爪变成红色,以便于雌性识别。它还以爪敲打胸腹板,或敲打步足以产生声音信号,其敲打次数及间隔长短也因种而异。这种敲打声在水中可传到50~100cm之外,而被同种雌性个体足部本体感受器所接受,以吸引雌性到雄性的穴中进行交配,并在此产卵孵化,直到幼体出来。雄性也可以直接敲打雌性。交配时一般雄性以第一对腹足插入雌性阴道,精子沿腹足中的沟进入雌体受精囊。如雌体是没有受精囊的种类,则雄性产生精荚,精荚附着在雌体最后两对胸足的基部,当雌性产卵时,由精荚释放出精子以受精,并在交配后立刻产卵。少数种类直接产卵于水中,例如对虾(Penaeus)。但大多数种类产卵于腹部附肢间,并有卵膜物质将卵粘附在一起。
多数种类于产卵时受精,受精卵靠附肢的打动造成水流以提供充足的氧。低等的甲壳类可形成卵袋,位于附肢两侧或身体背部,受精卵常变成桔红色。某些低等的甲壳类行完全卵裂,如鳃足类、桡足类表现出某些螺旋卵裂痕迹。高等的甲壳类均为中黄卵,行表面卵裂,具中空囊胚。发育中有自由游泳的幼虫,而且不同的种类有不同的幼虫类型。
对虾最早出现的幼虫是无节幼虫(nauplius),身体呈卵圆形,不分节,具三对附肢,第一对为单肢型,后两对为双肢型,相当于成体时的第一、二触角及大颚,具中眼一个。在发育中经连续蜕皮,躯干体节及相应的附肢逐渐形成,当前8对胸肢形成后称为溞状幼虫(zoea)。但大多数海产虾类初孵化的幼虫(第一期幼虫)为前溞状幼虫(protozoea)或溞状幼虫,其无节幼虫期在卵中度过。
当所有的附肢获得后称为后幼虫期(postlarva)。后幼虫期在外形上可能与成体相同,也可能不相同。例如蟹的后幼虫期腹部仍很长,位于体后并未折回到胸部下面。后幼虫经蜕皮后完全具备成虫特征,只是性器官未成熟。在此基础上,不同种类的甲壳类可有不同程度的变化。例如对虾的发育主要经过无节幼虫、前溞状幼虫、糠虾幼虫(相当于溞状幼虫)及长眼柄幼虫(后幼虫)。一些陆生或半陆生种类,繁殖时常返回淡水或海水中以完成其发育。
甲壳类的成虫仍然可以蜕皮,但蜕皮的次数少于幼虫期。少数种类成虫后不再蜕皮,例如蟹类。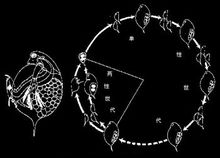
激素及其调节作用在无脊椎动物中甲壳纲与昆虫纲的激素及其调节作用是研究得最清楚的。这里所说的激素是指内激素(hormone)。外激素(pheromone),也称信息素,将在昆虫纲中介绍。激素是由内分泌腺体(endocrine gland)或神经分泌细胞(neurosecretory cells)所分泌的微量物质,直接送入血液或淋巴液,起调节动物生长发育的作用。
甲壳类的激素分泌腺体包括眼柄的视神经节内的几丛神经分泌细胞称为X器官,它分泌蜕皮抑制激素(moultinhibitinghormone)。X器官的分泌物通过轴突直接运送并贮存在窦腺(sinus gland)内,再由窦腺释放到血淋巴。另外,在眼柄的腹面及两侧还有神经分泌细胞,称Y器官,它分泌蜕皮激素(moult hormone),引起甲壳类的蜕皮。完成蜕皮后,X器官分泌的蜕皮抑制激素使之进入蜕皮间期。实验证明移走Y器官则阻止蜕皮,移走X器官则引起提前蜕皮。引起激素分泌的原因是由于外界刺激作用于中枢的结果,例如光照的长短、温度的变化等等。
甲壳类的生殖及第二性征也受激素调节。例如,在性成熟之前移走Y器官,则生殖腺的发育严重减缓;而在性成熟之后移走,则不受影响。卵巢也与X器官-窦腺系统之间存在着激素调节关系,非生殖期窦腺释放的激素抑制卵的发育。在生殖季节,中枢神经分泌一种促生殖腺激素,而血液中的生殖腺抑制激素水平下降,卵开始发育。卵巢本身也能产生某种激素,引起第二性征的出现,例如腹足上携卵刚毛的出现,卵袋的形成等。雄性精巢及第二性征的发育也被激素所控制,位于输精管末端附近的促雄性素腺(androgenic gland)(少数位于精巢内)所产生的激素具有这种作用。移走这一腺体雄性特征消失,且精巢转变成卵巢。如将这一腺体植入雌性,则卵巢变精巢,雄性性征又出现。
另外激素也控制着体壁色素细胞的变化,使体色适应外界环境,这在前面已述及。许多甲壳类表现出的某种生理节奏(physiological rhythmic)也是受激素控制的。例如一种绿泳蟹(Carcinus)及招潮蟹,都生活在潮间带,前者在高潮时活动,后者在低潮时活动,但两者都通过色素细胞的移动而使体色夜间变浅,白昼时变深暗,表现出色素细胞的节奏移动。甚至将它们饲养在连续光照或连续黑暗的条件下,这一节奏也不改变。但移走眼柄,这一节奏变化消失。相似的节奏如龙虾(Palinura)的昼夜活动,桡足类的昼夜垂直迁移都是由激素控制的。
自切与再生许多甲壳类,特别是蟹及寄居蟹等经常发生附肢的自切现象(autotomy)。自切的断面发生在附肢的基节与座节之间的一个双层膜处,膜上有孔,允许血管、神经通过,并由胸部体壁发出自切肌,直插入到断裂面。由于外力的牵引或自身因素,自切肌(autotomizer muscle)被刺激而剧烈收缩,由断裂面处分开,收缩的膜可堵塞血管以防止血液的外流。这种自切现象甚至可由单体节的反射活动而引起。
螯虾仅有螯肢能自切,寄居蟹前三对步足可自切,部分蟹五对步足均可自切。甲壳类也具有一定的再生能力。例如,断肢后可再生出新肢。之间需经多次蜕皮才能完成,再生部分可以达到丢失部分的大小。
演化如同其他节肢动物一样甲壳亚门的演化过程还不很清楚。这个不清楚的主要原因在于它们的甲壳比较难保存为化石。最早的甲壳亚门化石出现于寒武纪,发现的有介形亚纲和软甲亚纲动物的化石。最早的甲壳亚门动物有可能类似于生活在盐水洞穴中的桨足纲,但它们没有留下化石。鳃足纲出现于泥盆纪,蔓足亚纲出现于志留纪。
尤其介形亚纲动物的壳常常在沉积岩中出现,因此它是重要的指标化石。从它们出现以来它们就是浮游动物的重要组成部分。在化石中还常出现的有藤壶。
甲壳亚门的经济意义我国东南两面环海,海岸线长而曲折,生境复杂,浅海面积辽阔,内陆江河、湖泊纵贯星列,温度适宜,有利于甲壳动物栖息繁衍,孕育着丰富的甲壳类资源。其经济价值,首先是作为渔业的重要组成,在我国诸海产量最大的虾类是毛虾,其次是对虾。在黄渤海产的有白虾[Palae—mon(Exopalaemon)]和鹰爪虾(rrachypenaeus)'南海产的新对虾(Metapenaeus)、仿对虾(Parapenaeopsis)、鹰爪虾、赤虾(Metapenae—opsis)。南海产的龙虾和淡水产的沼虾,都是产量丰富的食用虾。蟹的种类也不少,海产产量最大的是三疣梭子蟹。虾蟹不仅味鲜美而且营养价值高,根据分析,河蟹的营养成分极为丰富,所含蛋白质与海蟹相等,较鲫为高,较沼虾、对虾、带鱼和鲤略低,所含脂肪和碳水化合物都特别丰富,远较上述几种食物为高。在矿物质方面,含铁质特别高,维生素A非常丰富,核黄素含量也多,硫胺素仅次于鲤、鲫。其次,甲壳动物又是鱼类的饵料,关系到天然鱼类的资源和养殖鱼类的繁殖和产量,我国沿海一带;进行对虾和中华绒螯蟹的人工饲养均已取得显著成效。然而甲壳动物也有对人类不利的一方面,如上所述,有些甲壳类是鱼类的寄生虫,严重的可导致幼鱼和戍鱼的大量死亡,有些甲壳类是人肺蛭的中间寄主,是该种寄生虫的传播媒介。海洋中的蔓足类附着于船舶、军舰外面,影响航速。蛀木水虱(Limnoria)和团水虱(卸haeroma)等钻蚀水下建筑材料,危害较大2。
生态与分布甲壳动物分布广泛,海洋、淡水和潮湿陆地上都可生存。大多数在海洋中,从潮间带(intertidal zone)直至万米深海中都能生活。许多种类营漂泊的浮游生活,如海水中的蜇水蚤、糠虾、嶙虾和毛虾等,淡水中的水蚤、剑水蚤、镖水蚤等。有的兼有游泳和爬行两种生活习性,如大多数虾类,虾蛄和能游泳的蟹等。有的在水底或岸上爬行,或常常潜居在泥沙洞穴中,碎石下,海藻间,如大多数蟹、跳钩虾等。还有一些种类是营固着生活的,如滕壶、茗荷儿等。有些甲壳动物与其他动物有着共栖、共生以及寄生关系。如双壳软体动物的外套腔内经常发现有成对的小虾,如江瑶虾、贝隐虾和小蟹如豆蟹,它们借贝壳以隐蔽,对贝类虽有一些影响,但并不危及其生存,是为共栖。还有附着在海绵、海葵、珊瑚、海参、海胆和海鞘等体外或体内(消循腔或体腔)。最有趣的是栖食在海绵动物“偕老同穴,,中的俪虾(spongicola venusta),它们在变态刚完成的幼虾阶段,便人海绵腔内,在其中长大直至老死,往往还是一雌一雄在内,故称俪虾。海葵和寄居蟹的共生,是大家十分熟悉的生态例子。
凡营寄生生活的甲壳动物,其形体多变异,它们多寄生在鱼体或其他无脊椎动物上,如中华鱼蚤、锚头鱼蚤等。寄生在蟹上的蟹奴(Sac,c以lina),附肢完全消失,只能从幼体辨别其为甲壳类。
此外,蟹有趋光性,游泳甲壳类随光线强弱而在水体作昼夜垂直分布的变迁,对虾由于气候、产卵等作长途洄游等,反映了甲壳动物生态的多样性2。
本词条内容贡献者为:
赵磊 - 副教授 - 中国农业大学
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国