

生物进化是指一切生命形态发生、发展的演变过程。“进化”一词来源于拉丁文evolution,原义为“展开”,一般用以指事物的逐渐变化、发展,由一种状态过渡到另一种状态。1762年,瑞士学者邦尼特最先将此词应用于生物学中。后来,达尔文的物种起源理论也是基于这一理论而诞生。
简介从遗传学角度来讲,睿智的先祖们,譬如达尔文、赫胥黎、海克尔及其他生物科学家,已经为我们确立了遗传法则在人类进化演变中所占据的主导性地位。人类的直立行走,以及其他种种生理能力运动、消化、血液循环、神经系统、肌肉力量、骨骼结构,甚至是精神能力,这一切都得益于人类的遗传。
尽管如此,还是遗漏了一种遗传,它超越了科学家们所能研究和想象的范畴。对于这种非凡的遗传现象的存在,他们深感无力无望,更是无法昭示于世人。只要大胆地敞开感官意识这扇大门,就能轻而易举地获得这股鲜活能量。那么,此时的你还犹豫什么呢?古人认为每件雕刻品都是种观念或情感的具体化,都是根据这样一个原则创作出来的,精神态与身体表达之间存在着完美的一致。而今我们承认精神状态与人的身体状况存在着直接的一致,而且只是一直是这样被阐述的,以至于我们如今知道,每一种境况都是结果,而这一结果,是一个源于某个理念的原因的产物1。
思想发展古代观念古代人们在栽培植物和驯养动物的生产实践中,积累了关于生物的形态、构造和生活习性的知识,注意到生物机体的变化以及生物与环境的关系,逐步形成了朴素的生物进化思想。古希腊的亚里士多德通过对他那个时代有关动物的知识的系统整理,把540种动物按性状的异同分为有血的和无血的两大群,每群之下又分为若干类。他进一步提出生物等级即生物阶梯的观念,认为自然界所有生物形成一个连续的系列,即从植物一直到人逐渐变得完善起来的直线系列;中国战国时期汇集的《尔雅》一书记载了生物类型的变化;汉初的《淮南子》一书,不仅对动植物作了初步分类,而且提出各类生物是由其原始类型发展而来的。
近代科学诞生以前,进化思想发展缓慢,当时广为流行的是神创论和物种不变论。这种观点直到18世纪仍在生物学中占统治地位,其代表人物是瑞典植物学家C.von林奈(1707~1778)。他所提出的分类系统虽然有助于揭示生物物种之间的历史联系,但他却把物种看作是上帝创造的不可改变的产物。随着生产和科学的发展,积累了许多新的与物种不变相矛盾的事实。在大量事实的影响下,甚至像林奈这样坚定的神创论者,在晚年也不得不承认由于杂交的结果能产生新种。
近代观念与林奈的观点相反,法国学者G.L.L.布丰(1707~1788)相信物种是变化的,现代的动物是少数原始类型的后代。他把有机体与居住环境联系起来,认为气候、食物和人的驯养等因素可引起动物性状的变异。1809年,另一位法国学者拉马克(1744~1829)在其《动物学哲学》中,用环境作用的影响、器官的用进废退和获得性的遗传等原理解释生物进化过程,创立了第一个比较严整的进化理论。其意义在于:否定了神创论和物种不变论,奠定了科学生物进化论的基础,即生物都不是神创造的,而是由更古老的生物进化而来的。
物种起源1859年,C.R.达尔文发表《物种起源》一书,论证了地球上现存的生物都由共同祖先发展而来,它们之间有亲缘关系,并提出自然选择学说以说明进化的原因,从而创立了科学的进化理论,揭示了生物发展的历史规律。
19世纪80年代以来,以A.魏斯曼(1834~1914)为代表的新达尔文主义把种质论和自然选择学说相结合,丰富了达尔文的进化理论。20世纪30年代以来,以T.杜布尚斯基(1906~1975)等人为代表的综合进化论综合了细胞遗传学、群体遗传学以及古生物学等学科的成就,进一步发展了以自然选择为核心的进化理论。60年代末,日本学者木村资生等人提出中性学说,又在分子水平上揭示了进化的某些特征,补充、丰富了进化论。
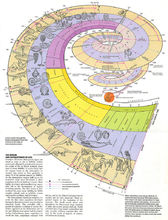
进化的进步性:地球上的生命,从最原始的无细胞结构状态进化为有细胞结构的原核生物,从原核生物进化为真核单细胞生物,然后按照不同方向发展,出现了真菌界、植物界和动物界。植物界从藻类到裸蕨植物再到蕨类植物、裸子植物,最后出现了被子植物。动物界从原始鞭毛虫到多细胞动物,从原始多细胞动物到出现脊索动物,进而演化出高等脊索动物──脊椎动物。脊椎动物中的鱼类又演化到两栖类再到爬行类,从中分化出哺乳类和鸟类,哺乳类中的一支进一步发展为高等智慧生物,这就是人2。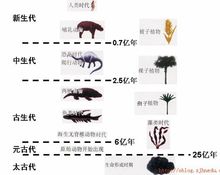
主要学说(1)拉马克的获得性状遗传学说
(2)达尔文的自然选择学说
(3)直生论
(4)生物进化的“突变论”
(5)新达尔文主义和综合进化学说
(6)中性学说
(7)“间断平衡”学说
(8)进化“四因说”
表1 主要进化理论的比较
|| ||
进化特征生物界的历史发展表明,生物进化是从水生到陆生、从简单到复杂、从低等到高等的过程,从中呈现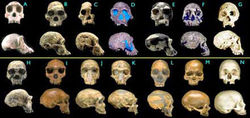 出一种进步性发展的趋势。
出一种进步性发展的趋势。
一般说来,进化过程的进步具有如下特征:
1. 在生物界的前进运动中,可以看到不同层次的形态结构的逐步复杂化和完善化;与此相应,生理功能也愈益专门化,效能亦逐步增高。
2. 从总体上看,遗传信息量随着生物的进化而逐步增加。
3. 内环境调控的不断完善及对环境分析能力和反应方式的发展,加强了机体对外界环境的自主性,扩大了活动范围。
生物进化的道路是曲折的,表现出种种特殊的复杂情况。除进步性发展外,生物界中还存在特化和退化现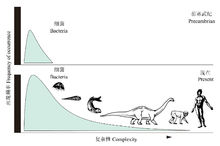 象。特化不同于全面的生物学的完善化,它是生物对某种环境条件的特异适应。这种进化方向有利于一个方面的发展却减少了其他方面的适应性,如马由多趾演变为适于奔跑的单蹄。当环境条件变化时,高度特化的生物类型往往由于不能适应而灭绝,如爱尔兰鹿,由于过分发达的角对生存弊多利少,以至终于灭绝2。对寄生或固着生活方式的适应,也可使机体某些器官和生理功能趋向退化。如有一种深海寄生鱼,雄体寄生在雌体上,雄体消化器官退化,唯有精巢特别膨大,以保证种族繁衍。
象。特化不同于全面的生物学的完善化,它是生物对某种环境条件的特异适应。这种进化方向有利于一个方面的发展却减少了其他方面的适应性,如马由多趾演变为适于奔跑的单蹄。当环境条件变化时,高度特化的生物类型往往由于不能适应而灭绝,如爱尔兰鹿,由于过分发达的角对生存弊多利少,以至终于灭绝2。对寄生或固着生活方式的适应,也可使机体某些器官和生理功能趋向退化。如有一种深海寄生鱼,雄体寄生在雌体上,雄体消化器官退化,唯有精巢特别膨大,以保证种族繁衍。
有些研究者对进化的进步性表示怀疑,认为进步性不是进化的基本特征,也不是进化的本质。科学研究证明,进化不全都引起进步,进化过程中也有退化,但从有机界总的进化过程看,进步性发展是进化的主流和本质。
进化方式 生物界各个物种和类群的进化,是通过不同方式进行的。物种形成(小进化)主要有两种方式:一种是渐进式形成,即由一个种逐渐演变为另一个或多个新种;另一种是爆发式形成,如多倍化种形成,这种方式在有性生殖的动物中很少发生,但在植物的进化中却相当普遍。世界上约有一半左右的植物种是通过染色体数目的突然改变而产生的多倍体。物类形成(大进化)常常表现为爆发式的进化过程(如寒武纪大爆发),从而使旧的类型和类群被迅速发展起来的新生的类型和类群所替代。
生物界各个物种和类群的进化,是通过不同方式进行的。物种形成(小进化)主要有两种方式:一种是渐进式形成,即由一个种逐渐演变为另一个或多个新种;另一种是爆发式形成,如多倍化种形成,这种方式在有性生殖的动物中很少发生,但在植物的进化中却相当普遍。世界上约有一半左右的植物种是通过染色体数目的突然改变而产生的多倍体。物类形成(大进化)常常表现为爆发式的进化过程(如寒武纪大爆发),从而使旧的类型和类群被迅速发展起来的新生的类型和类群所替代。
自然选择自然界的生物,通过激烈的生存斗争,适应者生存下来,不适应者被淘汰掉,这就是自然选择。
渐进式渐进进化是达尔文进化论的一个基本概念,达尔文认为,在生存斗争中,由适应的变异逐渐积累就会发展为显著的变异而导致新种的形成。因为“自然选择只能通过累积轻微的、连续的、有益的变异而发生作用,所以不能产生巨大的或突然的变化,它只能通过短且慢的步骤发生作用”。现代进化论坚持达尔文的渐变论思想和自然选择的创造性作用,强调进化是群体在长时期的遗传上的变化,认为通过突变(基因突变和染色体畸变)或遗传重组、选择、漂变、迁移和隔离等因素的作用,整个群体的基因组成就会发生变化,造成生殖隔离,演变为不同物种。20世纪70年代以来,一些古生物学者根据化石记录中显示出的进化间隙,提出间断平衡学说,代替传统的渐进观点。他们认为物种长期处于变化很小的静态平衡状态,由于某种原因,这种平衡会突然被打断,在较短时间内迅速成为新种。
爆发式对化石的研究发现,在进化史上,相当长的时间内处于进化较为沉寂的时期,新种的化石很少;有时大量的物种化石集中出现在较短的地质年代,如寒武纪大爆发。
寒武纪是古生代的第一个纪。研究证据表明,寒武纪大约是从距今5.44亿年前至距今5.05亿年前。地球上最早的生命大约是在距今38亿年前出现的。在从38亿年前到6亿年前这长达32亿年的时间里,生物进化的速率是十分缓慢的。最早的原核生物可能出现在35亿年前。最早的真核生物可能出现在20亿年前,从那时直至距今6亿年前,地球上的生物几乎都是单细胞生物。从寒武纪开始,地球上突然出现种类繁多的多细胞动物,人们称这种现象为“寒武纪大爆发(Cambrian explosion)”,也叫“寒武纪大爆炸”。
断续平衡论也称为间断平衡说。该学说认为化石的不连续性是历史的真实反映,这正说明生物的进化是不连续的,新物种是短时间内迅速出现的,然后是长时间的进化停滞,直到另一次快速的物种形成出现。
生物的进化既包含有缓慢的渐进,也包含有急剧的跃进;既是连续的,又是间断的。整个进化过程表现为渐进与跃进、连续与间断的辩证统一。
进化内容进化的基本单位种群是生物生存和生物进化的基本单位, 一个物种中的一个个体是不能长期生存的,物种长期生存的基本单位是种群。一个个体是不可能进化的,生物的进化是通过自然选择实现的,自然选择的对象不是个体而是一个群体。 种群也是生物繁殖的基本单位,种群内的个体不是机械地集合在一起,而是彼此可以交配,并通过繁殖将各自的基因传递给后代。
进化的原材料基因库和基因频率
基因库是指一个种群所含的全部基因。每个个体所含有的基因只是种群基因库中的一个组成部分。每个种群都有它独特的基因库,种群中的个体一代一代地死亡,但基因库却代代相传,并在传递过程中得到保持和发展。种群越大,基因库也越大,反之,种群越小基因库也越小。当种群变得很小时,就有可能失去遗传的多样性,从而失去了进化上的优势而逐渐被淘汰。
基因频率是指某种基因在某个种群中出现的比例。基因频率可用抽样调查的方法来获得。如果在种群足够大,没有基因突变,生存空间和食物都无限的条件下,即没有生存压力,种群内个体之间的交配又是随机的情况下,种群中的基因频率是不变的。但这种条件在自然状态下是不存在的,即使在实验条件下也很难做到。实际情况是由于存在基因突变、基因重组和自然选择等因素,种群的基因频率总是在不断变化的。这种基因频率变化的方向是由自然选择决定的。所以生物的进化实质上就是种群基因频率发生变化的过程。
基因频率的计算方法
设二倍体生物种群中的染色体的某一座位上有一对等位基因,记作A1和A2。假如种群中被调查的个体有N个,三种类型的基因组成,A1A1、A1A2和A2A2,在被调查对象中所占的个数分别为n1、n2和n3 基因库和基因频率的知识可与遗传的基本规律相结合,在深刻理解遗传的基本规律的基础上来理解基因库和基因频率的概念就容易得多,也很能够将这部分知识融会贯通。
基因频率的改变
引起基因频率改变的因素主要有三个:选择、遗传漂变和迁移。
选择即环境对变异的选择,即保存有利变异和淘汰不利变异的过程。选择的实质是定向地改变群体的基因频率。 选择是生物进化和物种形成的主导因素,已经发生的变异能否保留下来继续进化或成为新物种的基础必须经过自然选择的考验,则自然选择决定变异类型的生存或淘汰。自然选择只保留与环境相协调的变异类型(有利变异),可见自然选择是定向的。经过无数次选择,使一定区域某物种的有利变异的基因得到加强,不利变异的基因逐渐清除,从而改变了物种在同区域或不同区域内的基因频率(达尔文只是在个体水平上注意到不同性状的保留与否,而不能从分子水平对自然选择的结果加以分析),形成同一区域内物种的新类型或不同区域内同一物种的亚种,或经长期的选择,使基因频率的改变达到生殖隔离的程度,便形成新的物种。选择决定着不同类型变异的命运,也就决定了生物进化与物种形成的方向。
遗传漂变是指:如果种群太小,含有某基因的个体在种群中的数量又很少的情况下,可能会由于这个个体的突然死亡或没有交配而使这个基因在这个种群中消失的现象。一般而言,种群越小,遗传漂变就越显著。
迁移是指含有某种基因的个体在从一个地区迁移到另一个地区的机会不均等,而导致基因频率发生改变。如一对等位基因A和a,如果含有A基因的个体比含有a基因的个体更多地迁移到一个新的地区,那么在这个新地区建立的新种群的基因频率就发生了变化。
可遗传的变异
可遗传的变异是生物进化的原始材料,可遗传的变异主要来自基因突变、基因重组和染色体变异。在生物进化理论中,常将基因突变和染色体变异统称为突变。基因突变是指DNA分子结构的改变,即基因内部脱氧核苷酸的排列顺序发生改变。基因突变是普遍存在的。根据突变发生的条件可分为自然突变和诱发突变两类。不管在什么样的条件下发生突变,都是随机的,没有方向性。
染色体变异包括染色体结构的变异和染色体数量的变异,染色体数量的变异又包括个体染色体的增加或减少(非整倍数变化)和成倍地增加或减少(整倍数变化)两种类型。其中染色体结构的变异与非整倍数变异,由于破坏了生物体内遗传物质的平衡,所以一般对生物的生命活动是不利的,有时甚至是致命的,在生物进化过程中的意义不大。但染色体整倍数的变化没有破坏原有遗传物质的平衡,能够加强生物体的某些生命活动,对生物的进化,特别是某些新物种的形成有一定的意义如自然界中多倍物种的形成。
基因重组是指染色体间基因的交换和组合。是由于减数分裂过程中,同一个核内染色体复制后发生重组和互换,结果就产生了大量与亲本不同的基因组合的配子类型。又由于在有性生殖过程中,雌雄配子的结合是随机的,进一步增加了后代性状的变异类型。基因重组实际包括了基因的自由组合定律和基因的连锁与互换定律。
突变和基因重组都是不定向的,有有利的,也有不利的。但有利和不利不是绝对的,这要取决于环境条件。环境条件改变了,原先有利的变异可能变得不利,而原先不利的变异可能变得有利。等位基因是通过基因突变产生的,并在有性生殖过程中通过基因重组而形成多种多样的基因型,从而使种群出现大量的可遗传变异。 变异是不定向的,变异只是给生物进化提供原始材料,不能决定生物进化的方向1。
自然决定方向种群中产生的变异是不定向的,经过长期的自然选择,其中的不利变异被不断淘汰,有利变异则逐渐积累,从而使种群的基因频率发生定向的改变,导致生物朝着一定的方向缓慢地进化。
隔离产生新物种物种
物种是指分布在一定的自然区域,具有一定的形态结构和生理功能,而且在自然状态下能够相互交配和繁殖,能够产生出可育后代的一群生物个体。
隔离
隔离是指将一个种群分隔成许多个小种群,使彼此不能交配,这样不同的种群就会向不同的方向发展,就有可能形成不同的物种。隔离常有地理隔离和生殖隔离两种。
地理隔离是指分布在不同自然区域的种群,由于地理空间上的隔离即使彼此间无法相遇而不能进行基因交流。一定的地理隔离及相应区域的自然选择,可使分开的小种群朝着不同方向分化,形成各自的基因库和基因频率,产生同一物种的不同亚种。分类学上把只有地理隔离的同一物种的几个种群叫亚种。
生殖隔离是指种群间的个体不能自由交配,或者交配后不能产生出可育的后代的现象。一定的地理隔离有助于亚种的形成,进一步的地理隔离使它们的基因库和基因频率继续朝不同方向发展,形成更大的差异。把这样的群体和最初的种群放一起,将不发生基因交流,说明它们已经和原来的种群形成了生殖屏障,即生殖隔离。如果只有地理隔离,一旦发生某种地质变化,两个分开的小种群重新相遇,可以再融合在一起。
地理隔离是物种形成的量变阶段,生殖隔离是物种形成的质变时期。只有地理隔离而不形成生殖隔离,只能产生生物新类型或亚种,绝不可能产生新的物种。生殖隔离是物种形成的关键,是物种形成的最后阶段,是物种间真正的界线。生殖隔离保持了物种间的不可交配性,从而也保证了物种的相对稳定性。生殖隔离分受精前隔离和受精后隔离。教材中提到生物因求偶方式、繁殖期、开花季节、花形态等的不同而不能受精属于受精前生殖隔离。胚胎发育早期死亡或产生后代不属于受精后生殖隔离。
最新成果新的研究表明,生物在进化过程中,少数几代能够从人类导致的不利环境中生存下来,比如气候变化、过渡捕捞等等。这是因为生物会进化。
瑞典优密欧大学的生物学家托马斯-卡梅伦表示:“该发现颠覆了人们以前的观念,即进化的发生速度十分缓慢,往往需要数百甚至上千年的时间。实际上,进化速度很快,往往与生态变化联系在一起。”研究人员在实验室设定了杂乱的生活环境,并捕捉了一些野生土壤螨类,结果发现它们能迅速地适应生活环境。仅仅经过五代,这些野生螨就从基因层面进化出了新的特质,扭转了快要灭绝的势头。
卡梅伦表示:“那些具有基因选择性的螨类生长速度最为缓慢,但却拥有极强的繁殖力。因此,我们推测,从长期角度来看,该物种从衰退转变为兴盛的原因是环境对强繁殖力个体的自然选择作用,当然还包括对子代强适应力的选择作用。”
科技工作者经常使用螨类来研究广泛的生物学疑题,实验室的发现对人类捕猎动物的管理有着一定的指导作用,如对捕猎管理也许有必要将迅速的进化考虑进去2。
本词条内容贡献者为:
江松敏 - 副教授 - 复旦大学
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国