

生物固氮是指固氮微生物将大气中的氮气还原成氨的过程,固氮生物都属于个体微小的原核生物,所以,固氮生物又叫做固氮微生物。
根据固氮微生物的固氮特点以及与植物的关系,可以将它们分为自生固氮微生物、共生固氮微生物和联合固氮微生物三类。
简介生物固氮是根际生物对话的典型例证之一。自然界中的氮素资源十分丰富,大气中近80%的气体为氮素。但只有少数原核生物,即细菌和蓝绿藻(蓝藻细菌)能够固定空气中的氮素。这些原核生物通过自生或与植物共生,将大气中的氮气转化成能被植物吸收利用的氮素,称为生物固氮。其他的原核生物和真核生物均不能利用大气中的氮素。与工业固氮的高温高压条件相比,生物固氮在常温常压下就可以进行,是生物圈中氮循环的主要氮源之一,所固定的氮素在自然界中相当客观1。
分类生物固氮是指固氮微生物将大气中的氮还原成氨的过程。可以分为共生固氮微生物和自生固氮微生物两大类。
共生固氮微生物的特点是与一些绿色植物互利共生,如根瘤菌。它在土壤中分布广泛,呈棒形、“T”形或“Y”形,只有侵人到豆科植物的根内才能完成固氮作用,具有一定的专一性,某种特定的根瘤菌只能侵入某种特定的豆科植物(大豆根瘤菌只能侵入大豆的根,蚕豆根瘤菌可能侵入蚕豆、菜豆和豇豆)。它们与豆科植物的共生关系表现为:豆科植物通过光合作用制造的有机物,一部分提供给根瘤菌,根瘤菌通过固氮作用制造的氨则提供给豆科植物。其代谢类型为异养需氧型,而且固氮量较大。
自生固氮微生物的特点是在土壤中能够独立进行固氮的微生物,如圆褐固氮菌。它呈杆状或球状,它具有较强的固氮能力,并且能够分泌生长素,促进植物的生长和果实的发育。其代谢类型为异养需氧型,其固氮量较小2。
固氮形式联合固氮有些固氮微生物如固氮螺菌、雀稗固氮菌等。能够生活在玉米、雀稗、水稻和甘蔗等植物根内的皮层细胞之间。这些固氮微生物和共生的植物之间具有一定的专一性,但是不形成根瘤那样的特殊结构。这些微生物还能够自行固氮,它们的固氮特点介于自生固氮和共生固氮之间,这种固氮形式叫做联合固氮。
微生物在自然界中,有很多原核微生物,包括细菌和放线菌,它们可以在特定条件下把氮气还原为氨,因而被称为固氮微生物。固氮微生物的固氮过程完全是生物和微生物自发进行的,无须提供任何能源和设备,因而它减少了能源的消耗。由于全部固氮过程都是生物活动,无污染物排放,有利于保护生态环境。同时,由于减少和免除了化学氮素的投入,使农产品中硝酸和亚硝酸物质大幅度降低,提高了农产品的品质,减少致癌物质对人类的危害。

固氮菌原核微生物,属于自生固氮菌,其代谢类型是异养需氧型。利用的是土壤中的腐殖质,故在生态系统中的成分是分解者。
自生固氮微生物是指在土壤中能够独立进行固氮的微生物,其中,多数是一类叫做自生固氮菌的细菌。自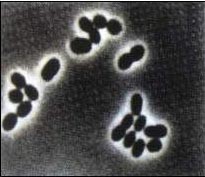 生固氮菌大多是杆菌或短杆菌,单生或对生。经过两三天的培养,成对的菌体呈“8”字形排列,并且外面有一层厚厚的荚膜。自生固氮菌中,人们应用得最多的是圆褐固氮菌(Azotobocter chroococcum)。圆褐固氮菌具有较强的固氮能力,并且能够分泌生长素,促进植株的生长和果实的发育,因此,将圆褐固氮菌制成菌剂,施用到土壤中,可以提高农作物的产量。
生固氮菌大多是杆菌或短杆菌,单生或对生。经过两三天的培养,成对的菌体呈“8”字形排列,并且外面有一层厚厚的荚膜。自生固氮菌中,人们应用得最多的是圆褐固氮菌(Azotobocter chroococcum)。圆褐固氮菌具有较强的固氮能力,并且能够分泌生长素,促进植株的生长和果实的发育,因此,将圆褐固氮菌制成菌剂,施用到土壤中,可以提高农作物的产量。
根瘤菌根瘤菌(root nodule bacteria)是与豆科植物共生,形成根瘤并固定空气中的氮气供植物营养的一类杆状细菌。这种共生体系具有很强的固氮能力。已知全世界豆科植物近两万种。根瘤菌是通过豆科植物根毛、侧根杈口(如花生)或其他部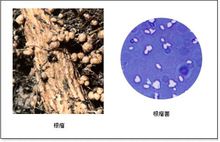 位侵入,形成侵入线,进到根的皮层,刺激宿主皮层细胞分裂,形成根瘤,根瘤菌从侵入线进到根瘤细胞,继续繁殖,根瘤中含有根瘤菌的细胞群构成含菌组织。
位侵入,形成侵入线,进到根的皮层,刺激宿主皮层细胞分裂,形成根瘤,根瘤菌从侵入线进到根瘤细胞,继续繁殖,根瘤中含有根瘤菌的细胞群构成含菌组织。
氮循环构成氮循环的主要环节是:生物体内有机氮的合成、氨化作用、硝化作用、反硝化作用和固氮作用。
植物吸收土壤中的铵盐和硝酸盐,进而将这些无机氮同化成植物体内的蛋白质等有机氮。动物直接或间接以植物为食物,将植物体内的有机氮同化成动物体内的有机氮。这一过程叫做生物体内有机氮的合成。动植物的遗体、排出物和残落物中的有机氮被微生物分解后形成氨,这一过程叫做氨化作用。在有氧的条件下,土壤中的氨或铵盐在硝化细菌的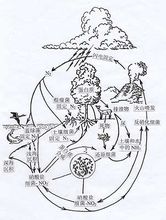 作用下最终氧化成硝酸盐,这一过程叫做硝化作用。氨化作用和硝化作用产生的无机氮,都能被植物吸收利用。在氧气不足的条件下,土壤中的硝酸盐被反硝化细菌等多种微生物还原成亚硝酸盐,并且进一步还原成分子态氮,分子态氮则返回到大气中,这一过程叫做反硝化作用。
作用下最终氧化成硝酸盐,这一过程叫做硝化作用。氨化作用和硝化作用产生的无机氮,都能被植物吸收利用。在氧气不足的条件下,土壤中的硝酸盐被反硝化细菌等多种微生物还原成亚硝酸盐,并且进一步还原成分子态氮,分子态氮则返回到大气中,这一过程叫做反硝化作用。
大气中的分子态氮被还原成氨,这一过程叫做固氮作用。没有固氮作用,大气中的分子态氮就不能被植物吸收利用。地球上固氮作用的途径有三种:生物固氮、工业固氮(用高温、高压和化学催化的方法,将氮转化成氨)和高能固氮(如闪电等高空瞬间放电所产生的高能,可以使空气中的氮与水中的氢结合,形成氨和硝酸,氨和硝酸则由雨水带到地面)。据科学家估算,每年生物固氮的总量占地球上固氮总量的90%左右,可见,生物固氮在地球的氮循环中具有十分重要的作用。
分子氮生物固氮是固氮微生物特有的一种生理功能,这种功能是在固氮酶的催化作用下进行的。固氮酶是一种能够将分子氮还原成氨的酶。固氮酶是由两种蛋白质组成的:一种含有铁,叫做铁蛋白,另一种含有铁和钼,叫做钼铁蛋白。只有铁蛋白和钼铁蛋白同时存在,固氮酶才具有固氮的作用。生物固氮过程可以用下面的反应式概括表示。
N2 + 6H+ + nMg-ATP +6e-(酶)→2NH3+nMg-ADP+nPi
分析上面的反应式可以看出,分子氮的还原过程是在固氮酶的催化作用下进行的。有三点需要说明:第一,ATP一定要与镁(Mg)结合,形成Mg-ATP复合物后才能起作用;第二,固氮酶具有底物多样性的特点,除了能够催化N2还原成NH3以外,还能催化乙炔还原成乙烯(固氮酶催化乙炔还原成乙烯的化学反应,常被科学家用来测定固氮酶的活性)等;第三,生物固氮过程中实际上还需要黄素氧还蛋白或铁氧还蛋白参与,这两种物质作为电子载体能够起到传递电子的作用。
铁蛋白与Mg-ATP结合以后,被黄素氧还蛋白或铁氧还蛋白还原,并与钼铁蛋白暂时结合以传递电子。铁蛋白每传递一个e-给钼铁蛋白, 同时伴随有两个Mg-ATP的水解。在这一催化反应中,铁蛋白反复氧化和还原,只有这样,e-和H+才能依次通过铁蛋白和钼铁蛋白,最终传递给N2和乙炔,使它们分别还原成NH3和乙烯。
1886年在第59届德国科学家和医生学术讨论会上,德国学者赫尔利格尔(Hermann Hellriegel)首次提出令人惊奇的试验结果,即当大豆生长在缺氮的土壤中时,大豆的根瘤也能使其良好生长,其机理在于其根瘤具有固氮功能。在当时称之为根生杆菌,现在称之为大豆根瘤菌的细菌对豆科植物根部的根瘤形成具有特殊的刺激作用。在根瘤菌内,根瘤菌将大气中的氮还原为能被植物吸收利用的氨,豆科宿主在吸收了这些氨之后又能将其转变为含氮有机化合物,以供其生长发育之需。通过对根瘤菌进行接种培养后于1895年就获得了具有很强固氮能力的根瘤菌菌种。通过添加灭菌草木灰等吸附剂之后,大批根瘤菌被施用到三叶草、豌豆和小扁豆等豆科作物的种植地中以提高其产量。
现在已经知道,在自然界具有固氮功能的生物种类很多,其中有自养固氮生物和异养固氮生物这两大类型。在异养固氮生物中因宿主植物的差异而被划分为豆科植物共生固氮菌和非豆科植物共生固氮菌。尽管固氮生物多种多样,但在其固氮过程中都需要共同的固氮基因(nif)的参与。在共生固氮生物中固氮体系非常复杂,除了nif基因在固氮过程中起关键性作用之外,其它基因的协同作用也非常重要。
由于根瘤菌具有的特殊功能,大批热心的研究者对其特征特性,对寄主的侵染方式、固氮机制和商业价值等进行了系统的研究。本世纪80年代以来,学者们一方面从分子水平进一步研究根瘤菌在豆科植物上的固氮机理和改造根瘤菌,试图培育出活性更强的根瘤菌;另一方面利用人工诱导方式诱发非豆科作物根部结瘤,试图利用根瘤菌的特殊功能使非豆科作物也能共生固氮,以便减少农田中氮肥的施用量,降低农作物的生产成本。除此之外,在70年代末,由于在放线菌中发现了弗兰克氏菌(Frankia)与多种非豆科树木能共生结瘤并具有固氮效应,因而在生物固氮研究中又产生了一个新的分支,即以研究弗兰克氏菌的分类、功能、分布和应用前景为主要内容的新领域。从现有的研究结果来看,与豆科植物的根瘤菌的固氮体系相比,利用弗兰克氏菌具有广谱侵染的特性,对建立新的固氮技术体系可能具有更大的意义,应用前景更广阔。
研究动态现在已经知道,生物固氮作用只限于原核类微生物(细菌和放线菌);所有不同种类的固氮微生物都由共同的固氮基因(nif)控制着固氮特性遗传,nif基因和固氮酶只存在于固氮菌体中;具有共生固氮特性的高等植物仅仅提供宿主条件,以便固氮菌的固氮效能得到充分表达;通过遗传操作实现了nif基因在不同种细菌之间相互转移,由此对nif基因的位置、数目、结构和功能等方面有了更深入的了解。
通过对自身固氮生物的nif基因进行分子生物学研究知道,在克氏肺炎杆菌Kp中存在着17~18个nif基因,这些基因都位于其染色体上,其中有固氮酶结构基因nifKDH、调节基因nifAL和固氮酶合成后的加工基因nifB以及其它与电子传递相关的基因。在同一固氮生物的个体中所有nif基因均连锁在一起,其中不存在非nif基因的插入成分;在nif基因群中存在着7个操纵子,其中有6个操纵子具有向同一方向进行转录的功能(Puhler,1984);在nifF基因和nifH基因之间还存在着一段反向阅读框架。
已经证实,在根瘤菌中的nif基因和结瘤基因都被定位在质粒上。根瘤菌的质粒有多种,其分子量为90×108或140×106或大于300×106。在根瘤菌的质粒中除了固氮基因之外还存在着结瘤基因(nod),使宿主的根毛变形弯曲的基因(hac)、根瘤起始基因(noi)和产生色素的基因(pig)等等。
在宿主植物中也有许多基因参与共生固氮作用,其中豆血红蛋白基因起着非常重要的作用。在豆科植物的根瘤内豆血红蛋白具有运氧功能,由此降低拟菌体周围的氧分压,以利于在嫌气条件下进行固氮作用。如果在根瘤内不存在着豆血红蛋白则为无效根瘤,这种根瘤不具有固氮功能。豆血红蛋白基因是隐性基因,只有当根瘤菌侵入宿主的根毛之后该基因才有可能表达。除此之外,在根瘤发育过程中还需要18~20种基因的产物(多肽)协同作用,这些基因统称为结瘤素基因,其多肽产物统称为结瘤素。
豆科植物固氮研究的发展趋势主要在两个方面,即应用研究和基础理论研究。在应用研究方面,试图培育出具有更强的固氮能力和具有吸氢酶基因的根瘤菌已经成为该领域的重要研究方向之一。霍尔斯特(Holster)等(1970)报道,根瘤菌可以与豆科植物在体外建立起共生关系,根瘤菌从细胞间隙进入大豆细胞内,但这种体外共生体系的固氮酶活性比较低,只相当于正常大豆根瘤的1%。通过研究工程化根瘤菌,筛选出新的固氮菌和根瘤菌接种剂,使豆科植物的结瘤数增加,固氮能力增强。已知固氮作用要消耗植物制造的光合产物的15~30%,在固氮过程中的放氢反应实际上是一种能量浪费。现在已经证实,在少数根瘤菌中存在着吸氢酶基因,其产物能回收在固氮作用过程中所放出的氢,由此形成ATP,节省能量,提高固氢效率。目前对这种吸氢酶基因已经进行了定位和克隆,如果能将这种基因转化到不具有吸氢酶基因的大多数根瘤菌中,则由此培育的新根瘤菌将具有更大的固氮活性和固氮效率。在基础理论研究方面,需要探明在根瘤菌与豆科植物共生固氮过程中双方有多少基因参与结瘤固氮作用,以便为建立人工模拟固氮体系提供理论依据;阐明根瘤菌对豆科植物专一性识别的机制,以便利用人工方法更精确地打破识别的专一性,扩大宿主范围;研究根瘤菌侵染宿主根毛之后,诱导宿主根内豆血红蛋白基因和结瘤素基因表达的机理,从而控制固氮根瘤菌的有效性。
非豆科在非豆科植物生物固氮的研究中主要包括两个研究方向。其一是采用适当的技术路线和切实可行的研究方法将根瘤菌导入非豆科作物中,在非豆科作物与根瘤菌之间形成共生关系,由此形成根瘤,扩大根瘤菌的宿主范围和提高其固氮效应。其二是采用先进的生物技术进一步对固氮放线菌Frankia的生物学特性及其应用价值进行探讨,以便挖掘Frankia菌在非豆科植物上的固氮潜力,由此形成生物固氮的新技术体系。
1.在非豆科植物与根瘤菌之间形成共生关系的探讨
德国植物学家拜尔(1888)首次在半寄生性草本植物草山萝和大猪鼻花(属于非豆科植物)的根部发现了根瘤,但这一奇特的现象并没有引起人们的关注。特里尼克(Trinick)(1973)首次证实,豇豆属植物根系中所存在的根瘤菌能与榆科植物共生结瘤固氮。帕甘(Pagan)等(1975)在试验中发现,在没有宿主植物细胞的情况下,豇豆根瘤菌能在人工培养基上独立生活和自行固氮,否定了长期以来一直认为根瘤脱离宿主植物就不能固氮的传统观念。如今已经知道,在残留的根部形成根瘤的非豆科植物的数量并不少,仅在俄罗斯的西伯利亚就有75个物种,分属于21个科,其中在进化史中最为年青的菊科植物中,其根系形成根瘤是一种最常见的现象。在新几内亚,在榆科的狗儿屎属植物Parasporiarogosa通常生长在茶叶树的行间,在其根部很容易发现与热带豆科植物相类似的结瘤现象。
在自然条件下植物体与微生物相结合后形成固氮体系的现象在70年代就已经引起了人们的关注。除了根瘤菌与豆科植物之间的共生固氮关系之外,还存在着兰绿藻与植物固氮;微生物与植物叶面结合固氮;土壤微生物与植物结合固氮;含脂刚螺菌与高等植物结合固氮。除此之外,对固氮稻的研究有了新的进展。在稻的根际嫌气性细菌和好气性细菌呈镶嵌状态,有着保护氧的作用,故稻的根际是最适于固氮菌存在的地方;现代栽培稻的固氮能力及各品种在固氮能力上所存在的差异是由其遗传性所支配,旱稻几乎没有固氮能力;在稻根际与固氮菌之间存在着松弛的共生关系。有关专家预测,通过寻找稻根际固氮基因和支配其固氮能力的基因,有可能利用具有更强固氮能力的基因或基因组,通过基因工程培育出固氮稻新品系。在80年代中期,将含有固氮调控基因nifA的质粒pMC78A引进到水稻根际固氮菌——催娩克氏菌(klebsiellaoxytocaNG18)中之后,筛选到耐氨菌株。将耐氨固氮菌接种到水稻根系之后,其根际固氮活性提高3~5倍,植株含氮总量提高20~30%,植株干重增加10~20%(日该国立遗传学研究所,1985~1986)。
自从聂延富(1983)利用植物生长素2,4-D处理根系,诱导根瘤菌侵入小麦根部形成根瘤的试验获得成功之后,中国在利用根瘤菌侵染非豆科植物形成根瘤方面开展了多方面的探索性研究,试图通过人工方法促使非豆科植物与根瘤菌形成共生关系,以便扩大生物固氮的范围。目前关于利用2,4-D诱导非豆科作物根系与固氮菌共生结瘤,国内外已在小麦、油菜、水稻和胡萝卜等作物上取得了成功。阿勒马拉(Al-Mallah)等(1989)利用纤维素酶yc、果胶酶Y23和甘露醇的细胞壁降解酶混合物处理幼苗根系,在有聚乙二醇存在的条件下将根瘤菌接种在水稻根系上,结果,在水稻根系上结出了根瘤。自80年代以来,中国相继对水稻、玉米、小麦和高梁等非豆科作物的联合结瘤固氮进行了深入的研究,成功地分离出一批具有高效固氮功能的联合固氮菌株,如粪产碱菌、稻黄杆菌和固氮螺菌等等。中国学者提出的“共生固氮体系中最佳结瘤固氮控制模型的研究”的探索性项目被列入国家的“攀登计划”。然而,总的来看,作为基础性研究,国内外在非豆科作物固氮方面虽不断取得新进展,但离农业生产的实际应用还有相当长的一段距离。
2.引导固氮生物侵入非豆科植物的方法
豆科植物的根瘤是一个非常复杂的共生体系,根瘤菌与豆科植物根系的相互结合依赖于特异的遗传机制。为了打破根瘤菌与豆科植物根系共生结瘤固氮的特异性,将豆科植物的共生固氮特性扩大到不具有共生固氮特性的非豆科植物中,研究者们从许多途径进行了有益的尝试,获得了一些有效方法,其中包括如下6种常用的方法。
第一,利用改造后的豆科植物根瘤菌直接侵染非豆科植物。利用基因工程技术构建携带有特定结瘤基因的质粒,然后将其转入到适当的根瘤菌中,由此获得发生质粒重组的受体菌,再利用受体菌直接侵染非豆科植物。
第二,利用植物裸露的原生质体吸收固氮微生物,以此建立起新的固氮体系。已经证实,裸露的原生质体比较容易吸收外源遗传物质,不但豆科植物的原生质体具有吸收固氮微生物的功能,而且非豆科植物的原生质体也具有吸收固氮微生物的功能。曾经利用玉米的原生质体吸收棕色固氮菌后,玉米原生质体表现出固氮活性,这样的原生质体可以培养成愈伤组织,由此分化出的胚在完全缺氮源的培养基上能够进一步生长发育。
第三,通过植物愈伤组织与固氮微生物进行联合培养,建立新的共生固氮体系。将豆科根瘤菌分别放入半支莲、胡萝卜、矮牵牛和囊龙面花等非豆科植物的愈伤组织中,经过联合培养后根瘤菌与愈伤组织形成新的共生关系,表现出固氮活性。
第四,利用酶处理打破根瘤菌侵染根毛的障碍。已知根瘤菌对宿主根毛区的侵染表现出很强的专一性,即特定的根瘤菌侵染特定的宿主。在玉米、水稻和小麦等作物上的试验结果表明,利用纤维素酶和果胶酶处理作物的根部后,其根毛细胞的细胞壁会发生局部破坏,这有助于根瘤菌的侵染和共生结瘤。
第五,利用植物激素打破根瘤菌侵染根毛的障碍。通过对非豆科植物的根系进行2,4-D处理后可以诱导豆科根瘤菌侵入其根皮层细胞内,随后可以形成根瘤菌。然而,更进一步的研究结果表明,在2,4-D诱导下在非豆科植物根系内所形成的根瘤与豆科植物的有效根瘤在结构上和功能上都存在着明显差异,其机理还有待于进一步研究。
第六,通过豆科植物与非豆科植物的原生质体融合后培育出新的固氮植物。在70年代中期将窄叶羽扇豆类菌体的原生质体与烟草叶肉原生质体融合成功后,获得了融合体。然而,目前在这方面的研究还有一些技术问题亟待解决。
3.在非豆科植物与Frankia菌之间形成共生关系的探讨
佩克洛(Peklo)(1910)首次提出从非豆科树木的根瘤中分离内生菌的设想,但他一直没有达到预期目的。在本世纪60年代,通过超显微观察后证实,在非豆科树木根瘤内所存在的固氮菌是原核生物,其丝状体有分枝。贝金(Becking)(1970)将非豆科树木根瘤内所存在的内生菌归属于原核生物界,厚壁菌门,放线菌纲,放线菌目,弗兰克氏菌科,弗兰克氏菌属。直到70年代末,卡拉哈姆(Callaham)等(1978)在香蕨木植物的根系中找到了共生根瘤并从共生根瘤中首次分离出内生放线菌。已经证实,在木麻黄、桤木、胡颓子、沙棘、杨梅、马桑、悬钩子、仙女木和赤杨等非豆科根瘤中所存在的内生固氮菌属于弗兰克氏(Frankia)菌属的微生物。近年的研究主要集中在对该菌的特征特性进行研究。由此发现,Frankia菌宿主范围比较宽,在非豆科树木的一些科、属、种间不仅存在交叉感染现象,而且由同一种树木的根瘤内可以获得在特征特性上存在着明显不同的Frankia菌株。Frankia菌是生长缓慢型微好气性放线菌,能形成特征性孢子囊,在孢子囊内含有通过纵横分裂所形成的不游动孢囊孢子。
在被子植物中能与Ftsnkia菌共生结瘤固氮的非豆科植物皆为乔木或灌木等木本植物,它们不仅具有强大的固氮能力,而且,通常还具有耐干旱(如沙棘)、耐盐碱(如木麻黄)、耐酸性(如杨梅)和耐水湿(如赤杨)等特性。据阿克曼(Akkermans)等人(1984)统计,已经发现21属200种非豆科植物能与Frankia菌共生结瘤固氮。随后,黄家彬等(1985)在中国又发现20种新记载树种具有与Frankia菌共生结瘤固氮的特性。最近,李志真等(1998)在福建省林区内发现5种能与Frankia菌共生结瘤固氮的非豆科树种。因此,目前已发现8科25属225种非豆科植物能与Frankia菌共生结瘤固氮。
中国于70年代末期开始研究放线菌结瘤植物,在资源调查、内生菌分离和Frankia菌的生物学特征特性等方面做了大量的研究工作。目前中国已经成为国际上拥有Frankia菌株资源最多的基地之一。除此之外,在利用细胞融合技术改进Frankia菌的特征特性方面已有新的苗头,利用链霉菌和Frankia菌融合后已经成功地构建成新型的共生固氮放线菌,它具有双亲本性能,即生长快、结瘤固氮活性强。
Frankia菌具有跨越科、属植物进行侵染结瘤固氮的特性,因而被认为是研究扩大寄主范围、结瘤机制、固氮基因转移和构建新的固氮物种的理想材料。随着对Frankia菌的特征特性的不断认识,人们试图将此共生体系的固氮功能由木本植物转到禾本科作物上,以解决农田氮肥自给问题,减少农作物对氮素化肥的依赖。随着研究的不断深入,人类将进一步揭示Frankia菌共生固氮的奥秘,阐明其遗传机制。利用生物工程技术和方法构建高效的Frankia工程菌和构建新的固氮共生体系将成为今后生物固氮研究领域中的发展方向和追求目标。
展望据测算,在大气中氮素含量为3.9×1015吨;在全球耕地内生物固氮量理论上可达到4400万吨,约相当于全世界每年生产的化肥总量;全球林地面积约为4.1亿公顷,其固氮总量可达到4000万吨。由于在氮素化肥生产中伴随着能源耗费和日趋严重的环境污染问题,人们逐渐认识到农林业生产完全依赖化肥终非良策,于是,生物固氮研究日益受到各国政府的重视。
通过适当方式固定大气中的游离氮素,将其转变为能参与生物体新陈代谢的氨态氮是地球上维持生产力的一个重要的生态反应。从战略上来考虑,正确的农业生产政策应该是既要增加粮食生产,又不要损害土地的持久生产力,而生物固氮正好能同时满足这两个目的。应用现代科学技术建立和完善生物固氮体系已经成为解决人类目前所面临的人口、粮食、能源和环境等问题的重要技术措施。
近20年来生物固氮已经成为一个多学科的综合性研究项目,分别在分子、细胞、个体和生态等多层次水平上,从微观到宏观不断地展开着探索性研究。从目前的研究现状来看,试图通过基因工程将nif基因从豆科植物转移到非豆科农作物中难度比较大,在短期内很难实现,而采用细胞工程方法将根瘤菌导入非宿主农作物细胞内则切实可行。除此之外,由于Frankia菌具有对宿主的侵染范围宽、固氮活性比较强和对氧气不敏感等特性,在生物固氮研究中对Frankia菌的研究将更为重要,有可能由此会找到新的突破口。在Frankia菌与农作物之间建立起新的共生固氮体系将具有更大的可能性。这项研究已呈现出新的苗头,值得进一步探索。
生物固氮研究已经引起越来越多的人的关注。在这方面的研究今后主要包括基础理论和应用基础这两个方面。在基础理论研究中主要围绕着诱发非豆科作物结瘤的最佳条件和提高共生固氮效能,其中包括诱导根瘤菌侵入主要农作物共生结瘤的有效方法;提高非豆科农作物共生结瘤固氮的效能;根瘤菌导入非豆科宿主细胞的途径、共生部位和共生机理;采用适当的技术措施诱导Frankia菌与主要农作物结瘤固氮;Frankia菌共生结瘤固氮的机理等等。在应用基础研究中主要围绕着培育新的固氮植物,其中包括通过生物技术改造固氮微生物和现有的农作物,使新的固氮菌与新的农作物更容易形成共生固氮关系。可以肯定,生物固氮工程的研究已经进入一个新的历史阶段,扩大生物间共生固氮范围和将豆科植物的固氮能力转移到非豆科植物中的研究已呈现出希望之光。随着生物固氮研究的不断深入,将逐步实现禾本科农作物与固氮微生物共生结瘤固氮的美好愿望。
农业应用简介生物固氮在农业生产中具有十分重要的作用。氮素是农作物从土壤中吸收的一种大量元素,土壤每年因此要失去大量的氮素。如果土壤每年得不到足够的氮素以弥补损失,土壤的含氮量就会下降。土壤可以通过两条途径获得氮素:一条是含氮肥料(包括氮素化肥和各种农家肥料)的施用;另一条是生物固氮。
拌种对豆科作物进行根瘤菌拌种,是提高豆科作物产量的一项有效措施。播种前, 将豆科作物的种子沾上与该种豆科作物相适应的根瘤菌,这显然有利于该种豆科作物结瘤固氮。特别是新开垦的农田和未种植过豆科作物的土壤中,根瘤菌很少,并且常常不能使豆科作物结瘤固氮,更需要进行根瘤菌拌种。对比实验表明,在其他条件相同的情况下,经过根瘤菌拌种的豆科作物,可以增产10%~20%。
将豆科作物的种子沾上与该种豆科作物相适应的根瘤菌,这显然有利于该种豆科作物结瘤固氮。特别是新开垦的农田和未种植过豆科作物的土壤中,根瘤菌很少,并且常常不能使豆科作物结瘤固氮,更需要进行根瘤菌拌种。对比实验表明,在其他条件相同的情况下,经过根瘤菌拌种的豆科作物,可以增产10%~20%。
做绿肥用豆科植物做绿肥,例如将田箐、苜蓿或紫云英等的新鲜植物直接耕埋或堆沤后施用到农田中,可以明显增加土壤中氮的含量。科学家统计过,如果用新鲜的豆科植物饲养家畜,再将家畜的粪便还田,则既可以使土壤肥沃,又可以获得更多的粮食和畜产品。
本世纪初以来全球农作物单位面积产量不断增长,在一定程度上依赖于氮素化肥的施用量不断增加。农作物依赖于施用氮素化肥所获得的增产实际上是以消耗能源和污染环境为代价所取得的。在大气中氮气含量接近80%,但这种氮气并不能直接为高等植物吸收利用。人类自从发现豆科植物与根瘤菌共生结瘤固氮现象以来对生物固氮研究已有112年之久,中国对生物的共生固氮现象也进行了长达62年的探索性研究。然而,关于生物固氮,特别是非豆科农作物的生物固氮,还有许多问题有待于进一步研究。目前,生物固氮研究已经被列为“国际生物学计划”中的重点研究内容,各国政府都将其视为重点科技攻关项目。通过适当方式将生物固氮机制引入到非豆科农作物中,进而建立起非豆科农作物固氮新体系,这是农业科学研究中一项富有挑战性的研究课题。这不仅引起了农业科学家的极大兴趣,而且也受到了全社会各阶层有识之士的广泛关注。
意义大气中的氮,必须通过以生物固氮为主的固氮作用,才能被植物吸收利用。动物直接或间接地以植物为食物。动物体内的一部分蛋白质在分解过程中产生的尿素等含氮废物,以及动植物遗体中的含氮物质,被土壤中的微生物分解后形成氨,氨经过土壤中的硝化细菌的作用,最终转化成硝酸盐,硝酸盐可以被植物吸收利用。在氧气不足的情况下,土壤中的另一些细菌可以将硝酸盐转化成亚硝酸盐并最终转化成氮气,氮气则返回到大气中。除了生物固氮以外,生产氮素化肥的工厂以及闪电等也可以固氮,但是,同生物固氮相比,它们所固定的氮素数量很少。可见,生物固氮在自然界氮循环中具有十分重要的作用。
本词条内容贡献者为:
李少斌 - 副教授 - 长江大学生命科学学院
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国