

γ-氨基丁酸(CAS号:1492-24-6 )别名4-氨基丁酸(γ-aminobutyric acid,简称GABA)相对分子量103.1,是一种四碳、非蛋白氨基酸,在脊椎动物、植物和微生物中广泛存在。1
GABA的简介物化性质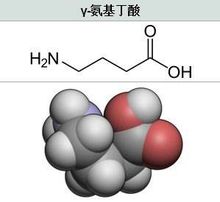 γ-氨基丁酸别名4-氨基丁酸(γ-aminobutyric acid,简称GABA),是一个四碳非蛋白质氨基酸,化学式:H2NCH2CH2CH2COOH;分子质量:103.1。GABA呈白色结晶体粉末状,没有旋光性。2熔点203℃(分解)34,与水混溶,微溶于乙醇、丙酮,不溶于苯、乙醚,分解时会失水生成吡咯烷酮。3其分子结构见右图。
γ-氨基丁酸别名4-氨基丁酸(γ-aminobutyric acid,简称GABA),是一个四碳非蛋白质氨基酸,化学式:H2NCH2CH2CH2COOH;分子质量:103.1。GABA呈白色结晶体粉末状,没有旋光性。2熔点203℃(分解)34,与水混溶,微溶于乙醇、丙酮,不溶于苯、乙醚,分解时会失水生成吡咯烷酮。3其分子结构见右图。
GABA在溶液中常以两性离子(带负电荷的羧基和带正电荷的氨基)形式存在,由于正负电荷基团间的静电相互作用,使得GABA在溶液中能够兼具气态(折叠态)和固态(伸展态)时的分子构象,而GABA在溶液中多分子构象共存的形式,使其能够结合多种受体蛋白并发挥多种重要生理功能。5
生理功能GABA作为一种抑制性神经递质起到分子信号的作用,具有多种生理功能。研究表明,GABA不仅具有降低神经元活性、防止神经细胞过热以及降低血压的作用,还具有防止动脉硬化、调节心律失常、降低血脂、增强肝功能等生理功效。而且,GABA对癫痫、惊厥、亨廷顿病和帕金森病等多种精神疾病具有一定的疗效作用。尽管 GABA具有重要的生理功能,但随着人们压力的增大和年龄的增长,人体中的GABA含量日益减少。因此,从食物中补充GABA对人体健康意义重大。
来源及应用植物组织中GABA的含量极低,通常在0.3~32.5μmol/g之间,不能满足人体的生理需求。已有文献报道,植物中GABA富集与植物所经历胁迫应激反应有关,在受到缺氧、热激、冷激、机械损伤、盐胁迫等胁迫压力时,会导致GABA的迅速积累。对植物性食品原料采用某种胁迫方式处理后,或通过微生物发酵作用使其体内GABA含量增加,用这种原料加工成富含GABA的功能产品已成为研究热点。GABA作为一种新型功能性因子,已被广泛应用于食品工业领域。利用富含GABA的发芽糙米、大豆和蚕豆等原料开发的食品已面市。1
允许添加剂量欧洲食品安全局(EFSA)虽然允许食物中添加GABA,规定GABA的膳食摄入量上限为550mg/d,但是其主要功能特性尚需严格的人群试验结果加以佐证。美国食品药品监督管理局(FDA)根据毒理学实验结果指出食品中添加GABA是安全的,使用范围包含饮料、咖啡、茶和口香糖等,但不允许在婴儿食品、肉制品或含肉产品中添加。中国卫生部2009年12号公告,GABA摄入量不得超过500mg/d,使用范围为饮料、可可制品、巧克力及其饮料、糖果、焙烤食品和膨化食品,但婴儿食品中不能添加。6
GABA的生物学功能GABA自动植物以及微生物中有较多的发现,其中在1949年首先在马铃薯的块茎中发现,在1950年又在哺乳 动物的中枢系统中发现其存在,同时被认为是哺乳动物、昆虫或者某些寄生蠕虫神经系统中的神经抑制剂,对神经元的兴奋程度有着重要的影响。2
降血压作用心血管方面的科学家通过研究发现对降低血压活性比较明显的红曲米和黄芪,他们的主要成分就是GABA。根据诸多的临床试验和动物实验,发现GABA对于降压和心脑血管调节有积极作用。2
调节心律失常包括人类的哺乳动物之中,都具有能够调节心血管以及心律的GABA系统,中枢GABA系统是必不可少的。引起心律失常的原因,就与中枢GABA系统受阻有关。有关研究表明,外源性GABA本身具有抑制心律失常的活性作用,并可以抵制试验性的心律失常行为。2
抗惊厥与抗焦虑作用GABA在人们的情绪调节上也有很大的作用,比如缓解人们的焦虑、抑郁等情绪的功能。这是因为GABA能够与延缓焦虑的脑部内受体进行两者的结合,同时作用于一些相关的物质,阻碍中枢神经的焦虑信息接收,从而达到缓解或者直接抑制焦虑的作用。几乎所有惊厥现象都和脑组织中GABA含量减少有关。抑制GABA合成的酶可减少GABA产生和阻碍GABA结合受体,使得抑制作用不能正常发挥,并诱发惊厥现象。相反,通过增加GABA降解酶的生成可以有效减少GABA的溶解,并增加它的存量,还可以通过服用地西泮类药物调节,使其与GABA的结合更加容易,对惊厥有较好的预防效果。2
调节激素的分泌大量的研究和试验验证表明,GABA能够通过下丘脑激素的释放来对GABA激导性进行传递,从而影响垂体分泌促性激素,GABA是脑内控制抑制性通路的主要物质,同时可以对内分泌系统的激素分泌状况进行干预,又可以促进腺垂体的生长激素分泌。所以提高血液中生长激素(GH、hGH)的含量可通过食用GABA实现。2
其他功能GABA在提神安宁以及提高睡眠质量和记忆力等生理层面也有比较突出的表现,包括对神经细胞以及脑细胞等,可以使它们延缓衰老,并且促使脑补血流顺畅、减缓喘息病症、对血压、脾脏、胃等器官都有良好的保护和营养改善作用,同时又可以帮助人们醒酒,更可以预防最常见的糖尿病,高血压等病症。2
GABA的制备方法1993有学者第一次通过化学合成的方法成功研制出了GABA。此后的相关研究日益丰富。GABA可开发作为具备显著药理功效的药物或保健食品,前景十分广阔。 为了获得更多的GABA,科研人员开始了各种尝试,并获得了诸多成果。2
化学合成法比较重要的化学合成主要有以下几种:第一种是采用邻苯二甲酰亚氨钾以及γ-氯丁氰或丁内酯作为制作GABA的原料,剧烈反应并水解后得到的最终产物就是GABA;第二种是利用吡咯烷酮作为最初的原料,并通过氢氧化钙以及碳酸氢铵进行水解,最终使其开环得到的产物就是GABA;第三种是把丁酸和氨水作为GABA的原料,使其在γ射线条件下进行光照反应得到GABA;第四种是通过辉光放电的方法,用丙胺和甲酸两种物质进行合成得到GABA;第五种是把溴醋酸甲酯和乙烯作为制备GABA的原料,通过聚合反应得到4-溴丁酸甲酯,最后经过氨解和水解后的产物即为GABA。GABA的化学合成方法都存在反应不容易控制、成本比较高的缺点。2
植物富集法植物富集法是一种新型开发的合成萃取提纯技术,它是用GABA含量较高的植物进行分离提取,这样便有了既便宜纯度又高的GABA产品。从植物中获取GABA的方法主要有以下两种:其中一种是利用溶剂萃取提纯法,另一种是柱分离制备法。2
(1)溶剂萃取法
溶剂萃取法是利用水或醇作为GABA的提取剂,根据植物在水或者醇中的溶解度以及分配系数不同的原理将GABA提出到水或者醇中,并且经过反复的过滤提纯,可以使植物中绝大多数的GABA都被萃取出来。2
(2)柱分离制备法
柱分离制备法,又被叫做柱色谱法,是一种利用不同的混合物中的组分在固液两相中具有不同分配系数的原理,进行洗脱分离及其他后续操作,它的大分类应该归属于层析法。色谱柱一般采用树脂、硅胶或活性炭等作为填充材料。2
微生物发酵法微生物发酵法是通过选择品种优良、稳定以及无毒无害的菌种,利用这些菌种在生长繁殖的过程中对GABA进行制备和产出。这种方法虽然对环境的要求比较苛刻,对设备的要求较高,但是此法产出的GABA可作为天然的食品添加剂。利用微生物发酵生产,是食品行业中发展最早,领域最广泛的生产方式之一,最早利用的微生物是大肠杆菌,利用它的脱羧酶可生产GABA,但是由于其本身存在一些安全隐患,使其一直无法直接用于药品或者食品的生产制作。2
随着科学技术的发展,绿色食品越来越受到人们的重视,后来科研人员发现乳酸菌、酵母菌以及曲霉菌等微生物都可以用来代替大肠杆菌,催化生产GABA。而且在较低成本的情况下,还具有产量高、安全性好的优势,此种方法已经逐渐在向产业化生产发展。2
植物中代谢途径在植物体中有两条GABA合成和转化途径:一条是谷氨酸经谷氨酸脱羧酶(glutamic acid decarboxylase,GAD)催化谷氨酸脱羧合成GABA,称为GABA支路(GABA shunt);另一条是由多胺降解产物转化形成GABA,称为多胺降解途径(polyamine degradation pathway)( 详见右图)1
GABA支路在高等植物中,GABA的代谢主要由三种酶参与完成,首先在GAD作用下,L-谷氨酸(glutamic acid,Glu)在α-位上发生不可逆脱羧反应生成GABA,然后在GABA转氨酶(GABA transaminase,GABA-T)催化下,GABA与丙酮酸和α-酮戊二酸反应生成琥珀酸半醛,最后经琥珀酸半醛脱氢酶(succinic semialdehyde dehydrogenase,SSADH)催化,琥珀酸半醛氧化脱氢形成琥珀酸最终进入三羧酸循环(krebs circle)。这条代谢途径构成了TCA循环的一条支路,称为GABA支路。1
在植物中,存在于细胞质中的GAD和线粒体中的GABA-T、SSADH共同调节GABA支路代谢,其中GAD是合成GABA的限速酶。植物GAD含有钙调蛋白(CaM)结合区,GAD活性不仅受Ca2+和H+浓度的共同调控,还受到GAD辅酶——磷酸吡哆醛(PLP)以及底物谷氨酸浓度的影响。这种双重调节机制将GABA的细胞积累与环境胁迫的性质和严重程度联系起来。冷激、热激、渗透胁迫和机械损伤均会提高细胞液中Ca2+浓度,Ca2+与CaM结合形成Ca2+/CaM复合体,在正常生理pH条件下能够刺激GAD基因表达,提高GAD活性;而酸性pH刺激GAD的出现是由于应激降低细胞的pH,减缓细胞受到酸性危害。植物中GABA支路被认为是合成GABA的主要途径。目前,大多数研究集中在如何提高GAD活性实现GABA富集。1
多胺降解途径多胺(polyamine,PAs)包括腐胺(putrescine,Put)、精胺(spermine,Spm)和亚精胺(spermidine,Spd),其中以腐胺作为多胺生物代谢的中心物质。多胺降解途径是指二胺或多胺(PAs)分别经二胺氧化酶(diamine oxidase,DAO)和多胺氧化酶(polyamine oxidase,PAO)催化产生4-氨基丁醛,再经4-氨基丁醛脱氢酶(4-amino aldehyde dehydrogenase,AMADH)脱氢生成GABA的过程,多胺降解途径最终与GABA支路交汇后参与TCA循环代谢。其中二胺氧化酶和多胺氧化酶是分别催化生物体内Put和Spd、Spm降解的关键酶。蚕豆发芽期间,厌氧胁迫可诱导多胺合成的关键性酶活性的提高,促进多胺的积累,同时多胺氧化酶活性也随之提高,通过多胺降解途径促进了GABA的合成与积累,提高了蚕豆的抗逆境能力。研究表明,大豆根中游离多胺含量在盐胁迫下增加,DAO活力提高,GABA富集量增加11~17倍。尽管多胺降解途径被认为是合成GABA的另一条重要途径,但其在单子叶植物中合成GABA的能力远低于GABA支路。1
微生物代谢途径在微生物中,GABA代谢是通过GABA支路完成的,利用微生物体内较高的GAD活性,将Glu脱羧形成 GABA,然后在GABA-T、SSADH作用下,GABA进入下游的分解过程生成琥珀酸半醛、琥珀酸参与微生物的生理代谢。微生物富集GABA就是通过对培养基的优化以及菌株的改良使其具有较高的GAD活性,增加GABA合成率,降低分解率来实现的。大量研究已证明GAD在原核到真核微生物中都有存在,此外,利用微生物中的GAD脱羧形成GABA不受资源、环境和空间的限制,与其他方法相比具有显著的优势。1
在高等动物中GABA的药用价值GABA具有特殊的药用价值,其对癫痫的治疗是近年来的热点之一。在几年来的抗神经性疼痛研究中发现大部分合成的GABA三唑衍生物具有抗癫痫、阵痛和抗痛觉过敏的作用。通过对癫痫发病患者和非癫痫患者的GAD 比对,发现癫痫患者GAD活性比非癫痫患者低。后来也有研究发现GABA的释放和GABA存储机制与癫痫存在密切关系。且GABA抑制作用减弱会导致癫痫发作。通过增加GABA代谢的药物对癫痫治疗已初见成效。目前癫痫发病原因被认为是神经自兴奋和抑制之间的平衡被破坏,而GABA在这其中起着重要的平衡作用。癫痫患者GAD神经元亚群易在发病中受损,导致GABA能神经元减少,而其他神经元则会上调GAD的表达以维持这种平衡。7
作为一个作用广泛的氨基酸,GABA还能影响癌细胞的增殖和凋亡、集落形成和侵袭,导致癌细胞有丝分裂活性变化,可作为治疗癌症的辅助药物。同样GABA可以提高自发性压力感受器反射功能(baroreceptor reflexfunction,BRS),调节心率降低血压。它还可以拮抗由咖啡因引起的睡眠障碍,且不影响自主活动。另外,也有研究发现GABA具有治疗糖尿病等作用。7
GABA有上述积极的作用同时,有时也不总是那么尽人意,甚至在动物体内还有许多不明确的生理和药理作用,需要更加谨慎的处理GABA和动物之间的关系。7
GABA的生物抗逆生理及调控作用GABA长久以来被认为与植物多种应激和防御系统有关。GABA会随着植物受到刺激而升高,被认为是植物中响应于各种外界变化、内部刺激和离子环境等因素如pH、温度、外部天敌刺激的一种有效机制。GABA还可以调节植物内环境如抗氧化、催熟、保鲜植物等作用。近年来GABA在植物中也被发现作为信号分子在植物中传递扩大信息。GABA曾在大豆、拟南芥、茉莉、草莓等植物中相继发现。低浓度的GABA有助于植物生长发育,高浓度下又会起相反的作用。7
GABA对外部酸化的响应低pH下GABA会在细胞内快速增加,这种GABA的积累在微生物和动物中也存在。植物在酸性pH下细胞内 H+随之升高,诱导细胞内GABA含量增加。该GABA的合成过程消耗H+,使得细胞内酸化得到缓解。在微生物中也存在这种快速的反应机制,在产生GABA的同时,会增加质子呼吸链复合物的表达,促进ATP合成。并且上调 F1F0-ATP水解酶活性,促进酸性条件下ATP依赖的H+排出过程。在动物中,细胞也会向外排出GABA和谷氨酸以此来改变细胞外环境的pH。更重要的是,GABA在生理环境下为两性离子,因此在酸碱调节中发挥着一定作用。7
GABA对昆虫的防御作用GABA有助于植物对外界天敌的防御。当昆虫取食时由于植物受伤导致细胞破裂和组织受伤,这种机械切割会刺激植物中Ca2+的增加,植物在Ca2+刺激下分泌GABA作为一种抵御昆虫取食的措施。在此过程中不存在茉莉酸类信号参与GABA的积累。昆虫存在离子型GABA受体,其中果蝇的GABA门控氯离子通道亚基RDL(resistant to dieldrin)是许多杀虫剂药物的作用靶标。GABA诱导使得GABA受体的单电流降低。具体为GABA在无脊椎动物中通过GABA受体门控的氯离子通道起作用,与大多数杀虫剂相同,通过GABA受体氯离子通道,使Cl-在电化学梯度的驱使下流向下游,导致质膜超极化,并抑制昆虫取食。而在过量表达GABA的烟草植物中,接种北方线虫,发现其雌性成年线虫的繁殖能力整体下降,这种方式可以使植物达到防御天敌的效果。在对女贞子被草食女娥幼虫取食过程中,发现女贞子会降低自身赖氨酸活性使得蛋白质无营养,而女娥幼虫在此期间会分泌甘氨酸、β-丙氨酸、胺等分子抑制植物赖氨酸的减少,这种植物与草食昆虫的交流过程也证明了GABA作为信号分子的功能。7
GABA对高等生物在高温和冷冻下的保护作用在小麦开花期间喷洒GABA(200 mg/L),可以调节膜稳定性,增加抗氧化能力等,减少了小麦高温下的损失;外源GABA的施用对黄瓜幼苗生长也有明显的作用。高温会抑制中枢GABA能神经元活性,激活胆碱类神经系统并引起体温升高。长期处于高温下,下丘脑的GABA能神经元活性会增加以适应环境和调节体温。GABA会在血浆中升高进而抑制冷敏神经核血浆中儿茶酚胺的浓度,达到降低食道温度的目的。7
低温会降低植物的生物合成能力,对重要功能造成干扰,并产生永久性伤害。动物在低温下也会导致损伤甚至造成更严重的伤害。低温下生物GABA表达会上调,这与低温的耐受性存在关联。在低温下,75%的代谢物会增加,包括氨基酸、糖类、抗坏血酸盐、腐胺和一些三羧酸循环中间体。能量代谢涉及的氨基酸代谢,酶类的转录丰度均会增加。可以通过增强GABA分流途径产生ATP以及积累GHB。另外低温下利用褪黑霉素可以使精胺、亚精胺和脯氨酸积累,促使二胺氧化酶表达升高。通过腐胺途径合成GABA,使得H2O2积累和苯丙烷途径通量下降以达到防腐和抗寒的效果。7
GABA在抗氧化和氧化过程中的作用GABA分流作为三羧酸循环分支途径的中间产物,与能量循环关系密切。同时GABA作为氧化代谢物的调控者发挥作用。将拟南芥SSADH突变体暴露于高温下生长,发现其活性氧中间体(reactive oxygen intermediate,ROI)积累,使得植株死亡,7证明ROI与GABA存在关系。同样SSADH和GABA-T基因的突变株在高温下存在大量的ROI,利用ROI消除剂N-叔丁基-α-苯基硝酮(PBN)可使GABA大量积累,从而提高酵母的存活率。因此,认为GABA分流途径在抑制高温下ROI具有作用。在GABA分流过程中,SSA可以经由GLYR/SSAR转化为GHB,而GHB与ROI存在密切关系。在SSADH缺失突变株中的GHB与ROI存在大量积累,而瓜巴特林可以抑制这种GHB与ROI的积累,并抑制了过氧化死亡。GABA分流过程可以减少ROI的积累使得生物免于高温带来的氧化损伤以及过氧化衰亡。7
GABA维持碳氮平衡碳氮代谢平衡涉及许多生理过程,包括能量代谢、氨基酸代谢等。由于GABA合成和分流途径涉及氮代谢,GABA也是能量循环中三羧酸循环的重要组成部分,GABA分流途径与呼吸链竞争SSADH,因此长时间以来 GABA被认为是碳氮代谢的重要一环。三羧酸循环分支的谷氨酸合成GABA途径是植物快速响应外部刺激的关键因素之一。绝大部分NH3+是通过谷氨酰胺合成酶/谷氨酸合成酶途径合成(glutamine synthetase/gluta-mate synthetase,GS/GOGAT),被认为是氨基酸的主要合成途径。游离的氨基分子大部分通过谷氨酰胺固定,谷氨酸被认为是植物老根中氮主要的积累形式,氮存储于精氨酸等氨基酸中,同时精氨酸也可用于运输,满足生物体的氮需求。同样氨基酸也通过转化为三羧酸循环的前体或中间体参与能量代谢过程。在对菠菜的研究中发现脯氨酸占总游离氨基酸的8.1%~36.%,GABA占12.8%~22.2%,谷氨酸占5.6% ~21.5%。谷氨酸是GABA和脯氨酸的前体物质,低温下植物会使谷氨酸的氮分流进入GABA和脯氨酸调控氮的代谢途径。另外在50mmol/L GABA下培养的拟南芥中除NADP+依赖性柠檬酸脱氢酶、根和芽中谷氨酰胺合成酶、芽中磷酸烯醇丙酮酸羧化酶外,几乎所有的初级氮代谢和硝酸盐吸收有关的酶活性均受到影响。而在NaCl条件下培养的拟南芥中,发现GABA积累的同时带动拟南芥整体氨基酸的增加。在分别利用不同氮化合物(10mmol/L NH4Cl,5mmol/L NH4NO3,5mmol/L谷氨酸和5mmol/L的谷氨酰胺)作为唯一氮源培养的拟南芥叶片中,其GAD活性和蛋白质水平不同,说明GAD在氮代谢中发挥作用。7
在NO胁迫下的香蕉中也发现了GAD活性上升、GABA和香蕉多巴胺增加的现象。盐胁迫下谷氨酸脱氢酶活性与GAD的表达瞬时上升,进而提高GABA分流等相关途径的通量以调节碳氮平衡。应激下NADH:NAD+和 ADP:ATP的比值也能影响GABA-T,从而使GABA积累。盐胁迫下植物更多地利用C/N平衡途径缓解压力。7
GABA在干旱和水涝中的作用20世纪末,人们就发现干旱可以降低根的固氮和O2的扩散,使得植物缺氧而导致GABA的积累。低氧条件下谷氨酸和天冬氨酸含量增加。干旱下GAD活性提高,GABA-T快速积累。干旱条件下,根系、茎的生长和叶面积伸展被抑制,活性氧增加,低分子渗透调节物质如GABA等氨基酸、多元醇、有机酸产量增加,以及抗氧化损伤的酶表达均上调。研究表明,干旱条件下,与细胞内稳态、活性氧的清除、结构蛋白稳定保护、渗透调节剂、转运蛋白等有关的基因表达上调。外源GABA使得植物保持较高的相对含水量,降低电解质渗漏、脂质、过氧化物、碳代谢并能提高膜稳定性。此外,外源GABA也可以诱导GABA-T和α-戊酸脱氢酶活性上升,抑制GAD活性使得GABA和谷氨酸增加。同时GABA加速多胺合成,抑制多胺分解,并进一步激活σ-1-吡咯林-5-羧酸合成酶和脯氨酸脱氢酶以及鸟氨酸-σ-氨基转移酶活性,致使GABA预富集物的高度积累和代谢。GABA还可以通过促进叶绿素表达,进而使得过氧化氢酶(catalase,CAT)、过氧化物酶(peroxidase,POX)活性增加,提高脯氨酸和糖含量,调节渗透和降低氧化。植物在水涝下pH会下降。长时间水涝会使土壤缺氧且短时间内水涝使得GABA升高。而水涝下气孔关闭与脱落酸存在直接关系。由于H+上升和缺氧会导致GABA增加。同时丙氨酸的积累可提高缺氧条件下植物的生存能力。在缺氧条件下GABA可以通过间接调节使得光合作用增强,降低气孔限制值,使得通氧量加大。缺氧条件下GAD活性上升,而GABA可以缓解缺氧对植物幼苗的伤害,而且外源GABA可以使低氧条件下根生长抑制得以缓解,快速生长出不定根。不定根生长也可以缓解植物的缺氧情况。7
另外,水涝缺氧条件下除GABA、谷氨酸以及丙氨酸外其他与三羧酸循环有关的氨基酸水平均下降。GABA与谷氨酸可作为丙氨酸的直接合成底物,通过这种厌氧途径生成2倍于糖酵解产生的ATP,保证供能。GABA还具有消除活性氧中间体以及为植物解毒和间接通过H2O2信号作用防止细胞程序性死亡(programmed cell eath,PCD),以及发挥其他作用。7
GABA的其他生理作用50mmol/L GABA和不同盐浓度会对植物幼苗产生不同的影响,当NO3-离子低于40mmol/L时,GABA会刺激 根伸长,当NO3-离子大于40mmol/L时GABA会抑制根伸长。并且GABA刺激低浓度的NO3-吸收,抑制高浓度NO3-的摄取,而GS等酶被氮调控,以上研究认为氮对调控植物生长有一定作用。在NaCl(50mmol/L)刺激下,植物的糖基化代谢会发起变化,并影响包括三羧酸循环、GABA代谢、氨基酸合成和莽草酸介导的次级代谢等发生变化。较高的盐离子会导致大豆的多胺氧化降解为GABA。植物GABA受体具有调节pH和Al3+的根耐受性。7
细菌侵染过程中的植物GAD表达量和γ-羟基丁酸转录丰度会上升,致使GABA升高。高GABA合成水平的烟草对根癌土壤杆菌C58感染敏感性有所下降。GABA可诱导农杆菌ATTKLM操纵子表达,使得N-(3-氧代辛酰基)高丝氨酸内酯的浓度减少,群体感应信号(或激素)下调,影响其对植物的毒性。GABA在植物与细菌的信号交流中也发挥作用,GABA可以抑制细菌内Hrpl基因表达(Hrpl基因编码蛋白使得植物致敏或引起其组织疾病),同时抑制植物体内hrp基因表达,使得植物免于过敏反应(hrp:控制植物病原体致病能力,并引起过敏反应)。7
此外,GABA还具有催熟作用。GABA可以通过刺激1-氨基环丙烷-1-羧酸(ACC)合成酶转录丰度刺激乙烯生物合成。而水涝下乙烯可以通过促进不定根的生长为植物提供氧气。高浓度GABA可抑制植物和细菌GABA转氨酶(GABA-T,GABT)突变体的生长,高浓度下可抑制细菌在植物内的繁殖。番茄中的GABA-T被抑制会导致GABA的积累,使番茄出现矮小症。7
展望在科研人员不断深入研究GABA下,了解到了GABA具有诸多药理活性。所以对于GABA的制备以及提取等研究内容越来越受到国内外科学家的重视,使GABA在食品领域的发展也成为一种重要的方向。2尤其是在日本,由于其成功研发出含有丰富的GABA的发芽糙米,其他相关含有GABA的产品相继被大力开发,并且受到人们的广泛好评。GABA现被人们广泛的开发用于食品、药品、保健品以及调味料等人们日常生活的用品中,并且可以适应不同的年龄以及人群结构。2
本词条内容贡献者为:
包申旭 - 教授 - 武汉理工大学

 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国