

出品;科普中国
作者:古明地恋(科普创作者)
监制:中国科普博览
提到海贝,你会想起什么?是碳烤生蚝、清蒸扇贝、辣炒蛏子,还是水煮毛蚶?
在大多数人的印象里,海贝的画面都与食物紧密相连,但这可不是因为你嘴馋,而是人类的共通印象。人类每年要吃掉超过1600万吨的软体动物,而其中70%都是双壳纲的贝类。
从以岸边贝类为食的东非早期人类开始,双壳贝类的肉就成为了人类的食物来源。它们过滤水流,将大量其他生物无法利用的浮游植物、浮游动物吸入体内,转化为营养丰富、细嫩脆爽的贝肉,再运送到人类的餐桌上。

肥美的牡蛎,壳中藏着满满的肉。
(图片来源:veer)
早期的腕足动物和双壳类动物
然而,人类能够吃到如此丰富的海贝,其实是一件十分幸运的事。在有生命出现以来的2亿多年的时间里,海洋中主要的滤食性动物都不是双壳纲的贝类,而是另一种叫作“腕足动物”的生物。
乍一看,腕足动物和双壳纲长得很像,都具有两瓣钙质壳,里面包藏着肉体,似乎吃哪种都相差不大,但它们的区别恰好就在肉体上。
双壳贝类壳内有着发达的软体,由外套膜包裹着丰富的肌肉、内脏和水管。外套膜的薄韧,闭壳肌的鲜甜,内脏的浓厚,水管的脆爽共同组成了贝类鲜美的滋味。
腕足动物却恰好相反,它们的壳很厚,内部肉却很少,一个不能吃的、坚硬的腕棒,加上其上生长的过滤卷须,就占了壳内的多达三分之二的内部空间,而肌肉、内脏则被挤到角落,可食部分的比例少的可怜。
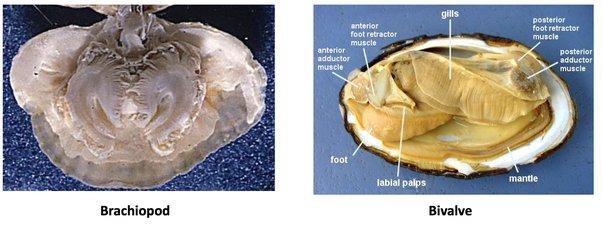
腕足动物(左)和双壳类(右)的比较,可见二者肉量的差异。注意,腕足动物身体中央占了最大比例的灰色腕冠和纤毛都是不能吃的
(图片来源:Quora)
海洋动物长肉,并不是专门给人吃的,而是为了自己的生活。肉体中含有具备生理活性的细胞,肉量越大,代谢速率就越快。肉质肥厚的双壳类,代谢速度相当于形容枯槁的腕足动物的3倍甚至10倍以上,能够产生更多的后代。

扇贝的肉体和壳
(图片来源:Wikipedia)
但比起双壳类,腕足动物占了“资历老”的优势。
早在5.4亿年前的寒武纪,滤食的腕足动物就已出现在地球上,它们在数亿年的时间里繁衍到了极多的数量,占据了先发优势。而在此时,早期的双壳纲还在像带着两瓣壳的小蜗牛一样钻在底泥里,以啃食底泥里的有机物质为生。当它们在4.8亿年前的奥陶纪终于演化出带有紧密鳃丝的合鳃,具备滤食能力的时候,海底的腕足动物早已繁殖到了极多的数量。
纵使肉质肥厚的双壳类代谢速度更快,在腕足动物压倒性的数量优势面前仍是不值一提。但这一状况,因为一场大灭绝事件而永久改变了。
地球生命灭绝中的地位转换
2.52亿年前的二叠纪末期,发生了地球生命最大的灭绝事件,西伯利亚地区的超级火山大规模喷发,巨量的岩浆吞噬了周边的一切生物,释放出的二氧化硫、烟尘带来的酸雨和寒冷严重破坏了海洋环境,全球冰封,海平面骤降;在二氧化硫的打击过去后,紧接着的二氧化碳又收割了最后一批生物。
在这次灭绝事件过后,双壳类的数量不久就超越了腕足动物,在世界各地都占据了优势,但腕足动物却再未恢复以往的繁盛,最终只能生活在阴暗寒冷的地区,成为海洋的边缘分子。
是什么原因导致了两者地位的转换呢?

模拟二叠纪末大灭绝的景象,火山大规模喷发
(图片来源:José-Luis Olivares/MIT)
自然而然的,人们想到了竞争。
腕足动物和双壳类动物具有相似的形态以及同样的生活方式,而双壳类在肉体方面有显著的优势。强大的肌肉允许它们进行挖掘、划水等运动,躲避敌害,寻找更好的食物来源;水管肌可激起强劲的水流,过滤更多的食物,获得更快的生长繁殖速率,产生更多后代。
也正因此,从19世纪60年代,也就是1860年左右开始,科学家们就假设两者具有竞争关系,具有生理优势的双壳类通过竞争,不断扩展分布,压缩腕足类的生存空间,逐步“消灭”了劣势的腕足类。
然而,这一基于猜想和“常理”的假设在最近受到了质疑。随着化石证据的增加,科学家可以从更加宏大的视角观察这个问题。
最近,中国地质大学(武汉)的博士生郭镇、陈中强教授和英国布里斯托大学教授迈克·本顿、博士生约瑟夫·费兰纳里-萨瑟兰合作,以庞大的古生物学数据库(PBDB)和数千枚化石为依托,重现了在二叠纪末大灭绝事件前后及后续演化中腕足动物和双壳类种类的变化。
研究表明,腕足动物在大灭绝事件中经受了严重的打击,而双壳类虽也有一定损失,但没有目以上的主要分类群的大规模灭绝,腕足动物比双壳类在这次大灭绝事件中受损严重得多。
大灭绝事件后,双壳类的物种数量立即开始恢复,在三叠纪最开始的一千万年里演化出大量的新物种,但与此同时,腕足类并没有一蹶不振,也同样生机勃勃地重新演化出新的类型。但在三叠纪开始200万年之后的奥伦尼克阶,腕足类又一次出现了大规模的灭绝,刚刚复苏的种群再次衰退,双壳类随即占据了绝对的优势。
然而,双壳类的优势并没有压制住腕足动物。它们在中三叠纪和中侏罗纪重新恢复,并在白垩纪的海洋中稳定地存在着,甚至在恐龙灭绝后的新生代进一步繁荣,在南大西洋西部热带大陆架的个别上升流环境中极其丰富,比双壳类和腹足类的总和还要多。
若是两者具有竞争关系,那么双壳类的多样化就会同时压制腕足类动物的恢复速度,直至其最终被大量的双壳类挤压空间而灭绝,但事实恰好相反。
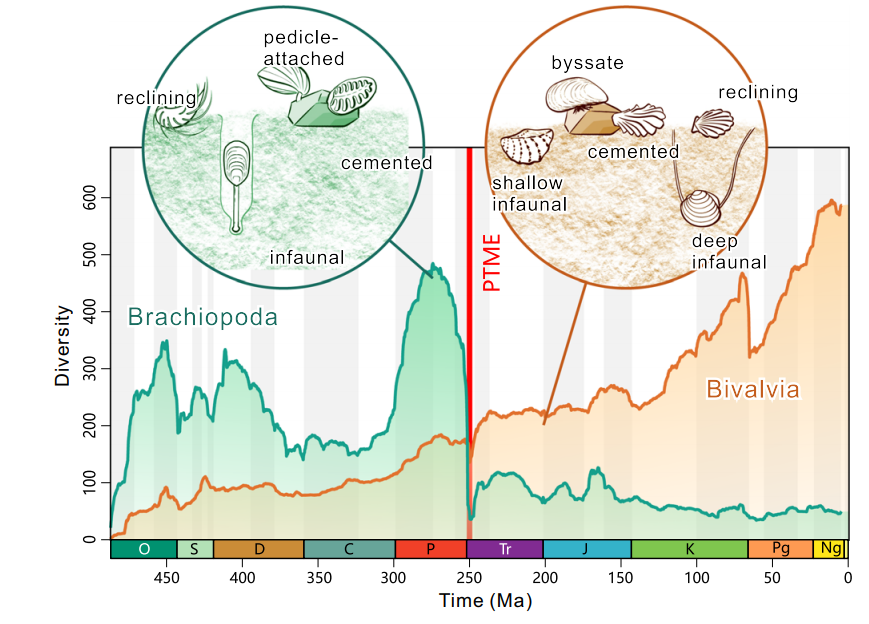
双壳类(橙色)和腕足动物(紫色)属数量的变化曲线,可看到腕足动物在二叠纪末受到了严重的打击,但在灭绝后仍有属数增加的时期,并非一路衰退。
(图片来源:参考文献[1])
抵抗大灭绝的生理优势
在大部分时间里,双壳类和腕足动物都是“一荣俱荣,一损俱损”的关系。二者需要同样的空间和食物资源,当环境改变时,双方自然会同时繁荣、衰退。而作为没有大脑、活动能力不强的滤食动物,两者虽有竞争,但无法像捕食者之间那样血雨腥风,更多的只是一些隐性的食物和空间的随缘争夺。
在平时,食物资源充足的大海完全容得下两种生物,双壳类和腕足类可以在不同的区域里生活,也可以混居在一起,一起享用洋流中的食物,几乎互不影响地各自占据着自己的海底份额。但是,如果碰到了大灭绝事件、海洋环境恶化的时候,那就只能各凭本事了。
很不幸的是,腕足动物肉少的生理劣势让它们难以抵抗大灭绝事件。大多数腕足动物完全是固着的,用肉茎或壳刺固定在沉积物或坚硬基质之上,进行不变位置的生活。

现代的腕足动物Liothyrella neozelandica,密集地分布在新西兰海面下的悬崖峭壁上
(图片来源:Ryan Photographic)
而肌肉发达的双壳类动物是可移动的“穴居专家”,它们主要生活在海床下方,挖掘不同深度的洞穴以保护自己,并演化出一种称为“水管”的吸管状结构,在埋在泥里时将食物和含氧的水带入它们的壳中。
另一些双壳类动物,尤其是扇贝,进化出通过贝柱(也就是闭壳肌)快速打开和关闭壳来游泳的方法,大大增加了它们逃避灾害和捕食者的能力。

鸟蛤肌肉发达的足部强壮而长,呈镰刀形,可以在沙中快速移动,躲避敌害和恶劣环境,同时也为它们带来了脆嫩、甘美,细嫩可口的味道。
(图片来源:Quora)
在大灭绝事件后不久的奥伦尼克阶,地球气温在强烈的温室效应下急剧升高约8—10摄氏度,延续近500万年之久,导致海水溶氧变得极其低下,大量的海洋生物都自此消失了。在其中,腕足动物受到的打击尤其严重。
说起原因,还是因为它们没肉。
厚壳阻碍了氧气的扩散,而小的肉体难以带动水流、加快呼吸,更无力推动自身移动,只能固定在原地窒息而亡。
相比之下,双壳类壳薄肉厚,强大的肌肉允许它们激起强劲的水流,在商场被贝类喷过水的朋友应该有体验;许多双壳类动物的壳上还有许多微细孔隙,能让水流通过,带来新鲜的氧气。在灾难下,这些能力便成为了它们少受低氧困扰的法宝。

埋栖在沙下的截形斧蛤,露出两只细长、肌肉质的水管,用来呼吸和过滤
(图片来源:Wikipedia)
而在同一时刻,腕足动物仍在与缺氧作斗争,不仅种群大量灭绝,幸存者也被迫减小体型,来适应低氧环境。对高温缺氧的不耐受最终造就了灾难性的后果:在这次大灭绝事件中,具有长浮游期幼体的腕足动物完全灭绝了,这几乎摧毁了它们重新广布地球的能力。
对于移动能力差的动物来说,长浮游期的幼体是底栖生物进行扩散的重中之重。它们能够在水中漂浮很长时间,通过洋流携带进行远距离的扩散,到达成体永远无法到达的遥远地方。
双壳类就具有这样的幼体,在一个月甚至超过一年的漂游期内,幼体最多可随水流飘出近4500千米,相当于横越四分之一个太平洋!这让双壳类能够在更多可生存的地区扎根,带来了它们广布全球的地理分布。
然而,二叠纪大灭绝事件后,幸存的钙质壳腕足动物只剩下了具备短浮游期幼体的类型。它们以自身携带的卵黄作为营养,不进食,只在水中飘散几天就沉降下来,固定在底质上便不再移动。较短的浮游期导致子代通常只能定居在距离亲代几千米以内,甚至与亲代紧密相连。
这种类型的优点是具有排他性——在一个狭小的区域内形成密集的群体,可以占据此地几乎所有的食物资源,排除其他滤食动物的竞争,但这也导致族群之间“内卷”严重,互相竞争食物资源,常常出现壳的畸形现象,亲代还会压迫子代使其窒息死亡。
更重要的是,幼体难以长距离扩散严重限制了它们的分布范围,每当新一轮灭绝到来,分布范围狭窄的腕足动物都有可能受到毁灭性的打击,两者此消彼长,造就了双壳类在以后的海洋中的主导地位。
在一次次的灭绝事件后,大部分的钙质壳腕足动物都只能生活在冷水和弱光的环境中,那里相对稳定、少受灭绝事件影响,但营养贫瘠。这意味着它们成为了海洋的边缘分子,几乎没有恢复从前繁盛的可能了。
与此同时,双壳类的数量则不断增长,遍布全球海洋的每个角落。它们丰润肥美,鲜嫩多汁的肉体帮助它们度过了无数次大灭绝事件,也让它们以最容易获得的肉类来源的姿态,养育了一代又一代的人类。

炒花蛤,我国最常见、产量最大的贝类。
(图片来源:veer)
结语
从世界各地的海岸线和文明遗址中发现的贝壳,到罗马人手中发光的海笋,《我的叔叔于勒》中文雅吸食的牡蛎,再到现代文明海岸线上连片的养殖场,无数的双壳类在历史上扮演着重要的角色,在数万年的时间里向全世界提供了无数的食品、贝壳和珍珠。而如果没有这次大灭绝事件,这一切都将不复存在。
所以你看,这次的大灭绝事件对当时的生物来说是灭顶之灾,对我们人类来说却不一定是坏事,自然多么奇妙啊。
参考文献:
[1]Guo, Z., Flannery-Sutherland, J.T., Benton, M.J. et al. Bayesian analyses indicate bivalves did not drive the downfall of brachiopods following the Permian-Triassic mass extinction. Nat Commun 14, 5566 (2023).
[2]Sun Y, Joachimski M M, Wignall P B, et al. Lethally hot temperatures during the Early Triassic greenhouse[J]. Science, 2012, 338(6105): 366-370.
[3]Gould S J, Calloway C B. Clams and brachiopods—ships that pass in the night[J]. Paleobiology, 1980, 6(4): 383-396.
[4]Liow L H, Reitan T, Harnik P G. Ecological interactions on macroevolutionary time scales: clams and brachiopods are more than ships that pass in the night[J]. Ecology letters, 2015, 18(10): 1030-1039.
[5]Kowalewski M, Simões M G, Carroll M, et al. Abundant brachiopods on a tropical, upwelling-influenced shelf (Southeast Brazilian Bight, South Atlantic)[J]. Palaios, 2002, 17(3): 277-286.
[6]Kowalewski, M., Hoffmeister, A. P., Baumiller, T. K. & Bambach, R. K. Secondary evolutionary escalation between brachiopods and enemies of other prey. Science 308, 1774–1777 (2005).
[7]Valentine J W, Jablonski D. Larval adaptations and patterns of brachiopod diversity in space and time[J]. Evolution, 1983: 1052-1061.
[8]Stanley S M. What has happened to the articulate brachiopods[C]//Geological Society of America Abstracts with Programs. 1974, 6(7): 966-967.
[9]Rhodes M C, Thompson R J. Comparative physiology of suspension-feeding in living brachiopods and bivalves: evolutionary implications[J]. Paleobiology, 1993, 19(3): 322-334.


 2023-09-26
2023-09-26


 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国