

据世界自然基金会统计,1970-2016 年,全球代表物种种群数量减少了 68%,生物多样性不断下降。
保护生物多样性,需要对当地生态情况进行准确分析,制定合理的生态保护政策。然而,生态数据太过庞杂,统计标准又难以统一,大规模的生态分析很难开展。
近期,康奈尔大学的研究者们利用深度学习,分析了 900 万组鸟类数据,得到了林莺在北美洲的分布数据,开启了生态数据分析的新篇章。
作者 | 雪菜
编辑 | 三羊、铁塔
本文首发自 HyperAI 超神经微信公众平台~
据世界自然基金会 (WWF) 统计,1970 年至 2016 年,全球 4,392 个代表物种、20,811 个种群的平均数量降低了 68%,全球生物多样性正在下降。

图 1:1970-2016 年,全球 4,392 个代表物种、20,811 个种群的平均数量变化
保护生物多样性,需要对相关地区的物种分布进行准确的大规模分析。然而,由于数据量过于庞大,且缺乏统一的统计方法,研究者们暂无法准确统计特定区域的生物多样性(物种丰富度、种群数量等)和生物组成数据(在当地生态系统中某一个物种的地位)。
传统的物种丰富度统计,需要将不同物种的分布地图叠加,进行建模预测,或是直接通过宏观生态学模型进行预测。无论哪种方法,推断结果都会受到模型精度的影响,前者还会受到地图精度的影响。
而且,这种预测方法的时间分辨率很差,无法对物种分布的季节性变化作出准确判断,更无法对物种间的联系进行研究,不利于生态保护政策的制定。
深度学习为生物多样性的大规模时空研究提供了有效手段。美国康奈尔大学的研究者们结合深度推理网络 (DRN, Deep Reasoning Network) 和深度多元 Probit 模型 (DMVP, Deep Multivariate Probit Model) 开发了 DMVP-DRNets 模型,从 9,206,241 组 eBird 数据中分析出了林莺 (Warbler) 在北美洲的时空分布,并对林莺与环境、其他物种之间的联系作出了推断。相关成果已发表于「Ecology」。

这一成果已发表于 「Ecology」
实验过程
数据集:eBird 与协变量
研究人员使用 2004 年 1 月 1 日至 2019 年 2 月 2 日, 170°-60° W,20°-60° N 之间的 eBird 数据作为本研究的数据集。排除重复数据后,共有 9,206,241 组 eBird 数据,每组 eBird 数据包括时间、日期、地点及观察到的所有鸟类物种。

图 2:一组银喉长尾山雀的 eBird 数据
研究人员还引入了 72 个协变量,包括 5 个与观察者相关的协变量,如活动状态、观察人数、观察时间等;3 个与时间相关的协变量,主要用于弥合不同时区之间的偏差;64 个与地形地貌相关的变量,如海拔、海岸线、岛屿等。
模型框架:解码器 + 潜在空间
本研究使用基于 DMVP 的 DRN 进行数据分析和预测。这一模型包含 3 层全连接 (fully-connected) 的网络解码器,用于分析输入特征的相关性,还有两个结构化潜在空间 (structured latent space) ,用以表示物种之间和物种-环境间的关联。

图 3:DMVP-DRNets 模型结果示意图
最终,DMVP-DRNets 模型通过一个可解释的潜在空间,输出 3 个生态相关的结果:
1、环境相关特征:反映了不同环境协变量之间的联系和相互作用;
2、物种相关特征:通过残差相关矩阵反映不同物种间的联系;
3、生物多样性相关特征:如某一物种的丰度和分布等。
模型评估:与 HLR-S 对比
将 DMVP-DRNets 模型投入大规模使用前,研究人员首先将其与基于空间高斯过程的 HLR-S 模型进行了对比。HLR-S 是生态学中研究多物种联合分布最常用的模型之一。
首先用 10,000 组 eBird 数据对两个模型进行训练。HLR-S 模型训练用时超过 24 小时,而 DMVP-DRNets 模型耗时不足 1 分钟。

表 1:DMVP-DRNets 模型和 HLR-S 模型性能对比
随后,对不同规模的 eBird 数据进行分析,DMVP-DRNets 模型在 11 个评价标准中优于 HLR-S 模型,仅在物种丰富度校准损失中落后于 HLR-S 模型。
实验结果
分布区域:阿巴拉契亚山脉
在对 eBird 的数据进行分析后,DMVP-DRNets 模型输出了空间分辨率为 2.9 km2 的北美林莺各月分布图。不同品种林莺在北美的分布动态性很强,每个月都有不同的分布热点。在对各月分布图进行叠加后,研究人员发现阿巴拉契亚山脉是林莺物种多样性最高的区域。
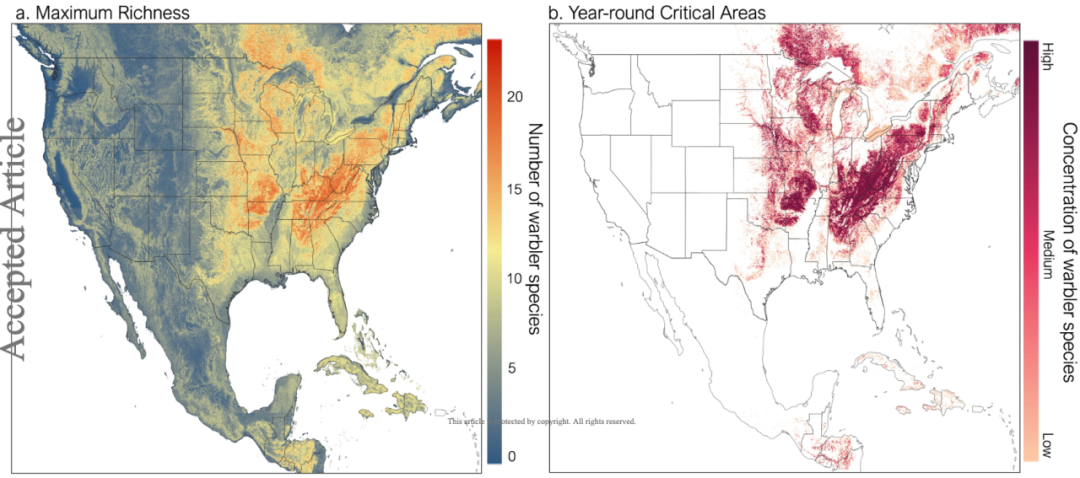
图 4:北美洲的林莺分布图
a:林莺在北美洲各地的最大物种丰富度分布
b:林莺在北美洲的主要分布区域
同时,研究人员还发现了不同迁徙期的林莺分布热点。在繁殖前迁徙期,林莺主要分布在俄亥俄州、西弗吉尼亚州和宾夕法尼亚州的阿巴拉契亚山脉附近。而在繁殖后,北阿巴拉契亚山脉是林莺分布最多的区域。
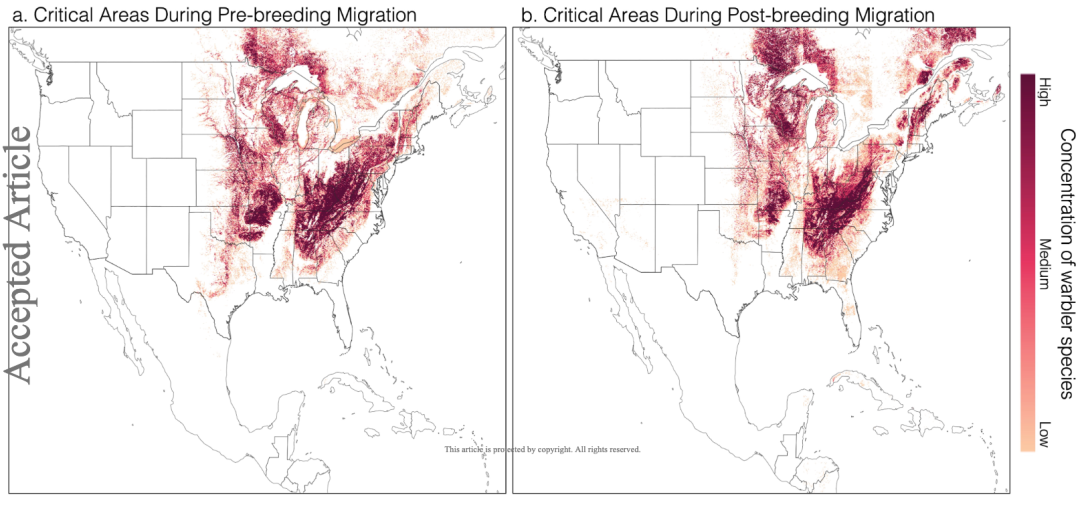
图 5:繁殖前迁徙期 (a) 和繁殖后迁徙期 (b) 的林莺分布
林莺-环境:水陆与季节偏好
进一步的,研究人员利用 DMVP-DRNets 模型对美国东北部的林莺-环境间的相互作用进行了分析。
首先,研究人员能够大致分辨出不同林莺对水生环境和陆地环境的偏好。随后,他们发现在繁殖期不同品种林莺对于环境的喜好不同。喜好水生环境的蓝翅黄森莺、北森莺和黄喉林莺在繁殖期栖息较近,而松林莺会和其他与松林相关的物种走得更近,如棕头鳾和红头啄木鸟。
随着季节的变化,不同林莺的分布也有变化。在繁殖后迁徙期,大多数林莺会抱团栖息,而棕榈林莺会选择在稍晚的秋天迁徙。松林莺和黄腰白喉林莺则会整年栖息在美国东北部。
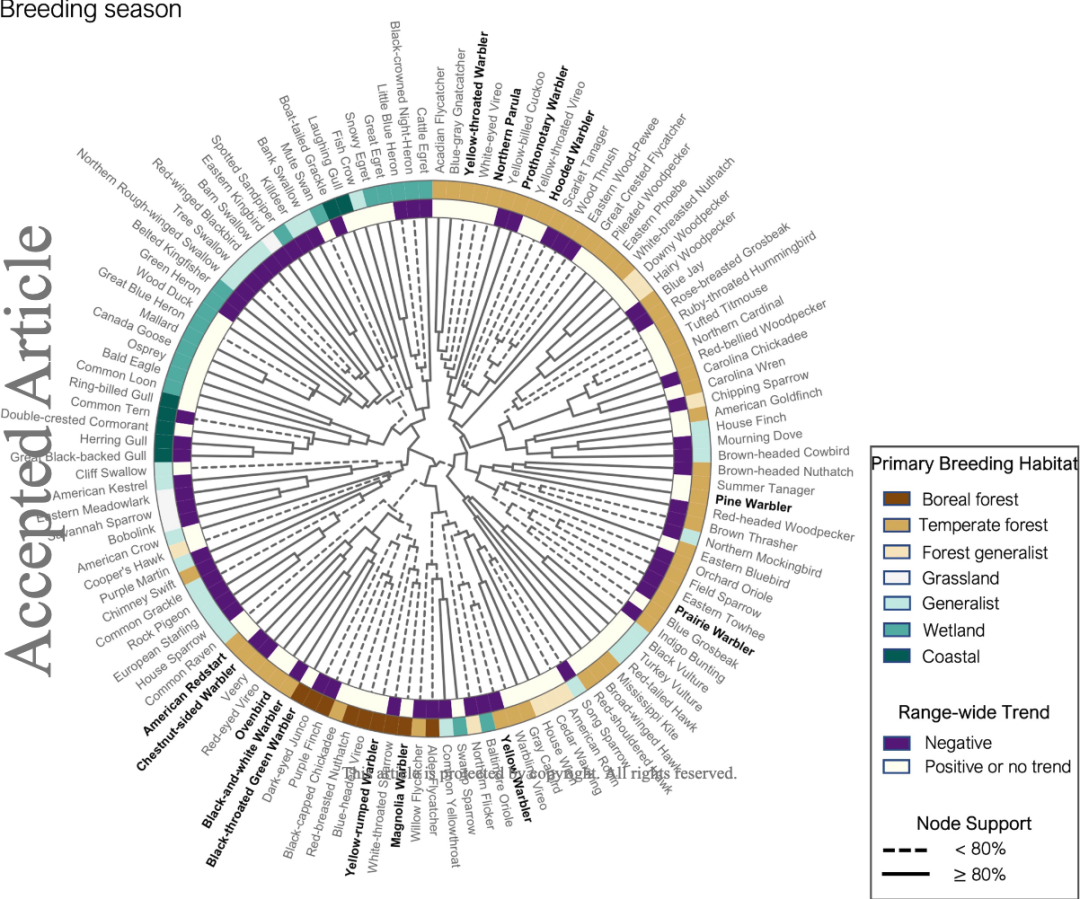
图 6:繁殖期林莺与环境、其他物种之间的关联性
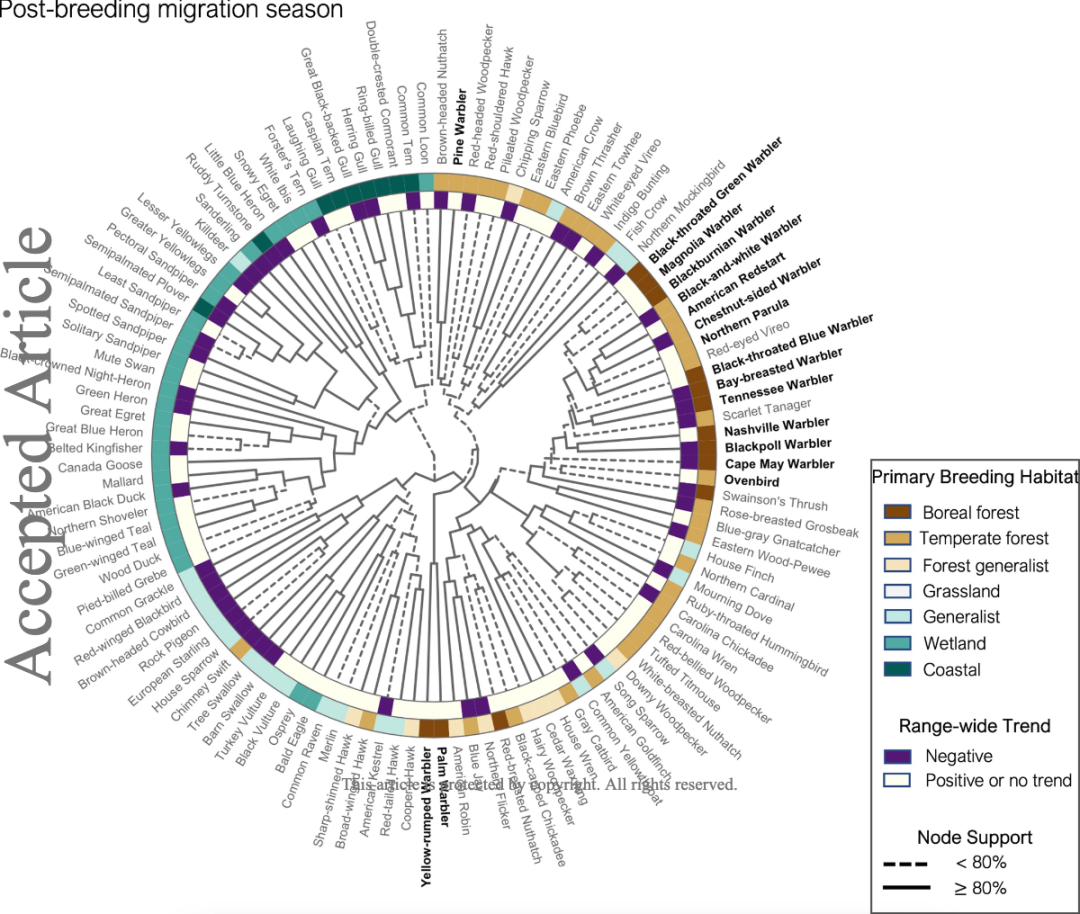
图 7:繁殖后迁徙期林莺与环境、其他物种之间的关联性
物种间关联:竞争与合作
在繁殖期、非繁殖期和迁徙期,林莺与其他物种之间展现出了不同的关系。
在繁殖期,林莺主要在防御自己的栖息地,与其他物种关联较弱。在栖息地相近且进攻性较强的品种之间,甚至是负关联,如黑枕威森莺和橙尾鸲莺。
在迁徙期,大多数林莺之间展现出了较强的正相关,与森林中的其他物种也有强关联性。这与观察的结果一致,林莺会与红眼绿鹃、黑冕山雀等其他物种组成混合迁徙队伍。
在此期间,林莺与巨翅鵟、条纹鹰、鸡鹰、赤肩鵟等捕食者关系较差,二者负相关系数较高。
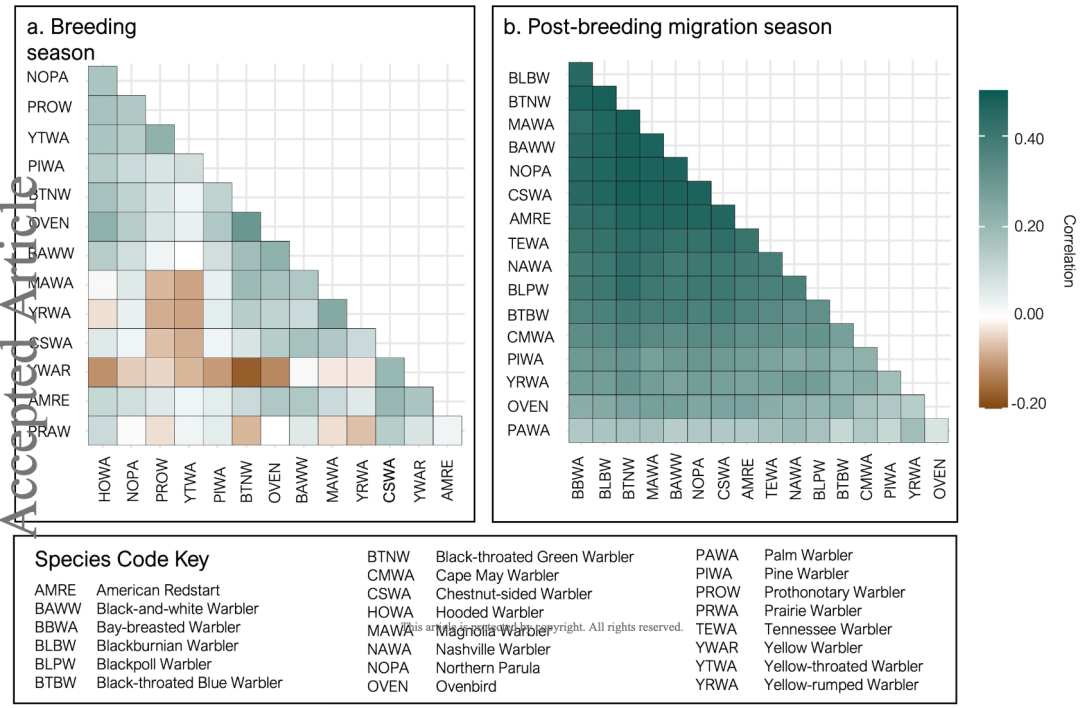
图 8:繁殖期 (a) 和繁殖后迁徙期 (b) 林莺与其他物种的相关系数
上述结果说明,DMVP-DRNets 模型可以对不同时期的林莺分布作出准确的判断,并能够推断出林莺与环境、其他物种之间的联系,为制定生态政策提供依据。
AI 「鸟口普查」
除了数据分析,数据采集也是生态研究的重要部分。与植物不同,鸟类的警觉性很高,动作迅速,且有些品种个体较小,很难进行准确观测。
传统方法依赖长焦相机、高倍望远镜和静止摄像头从远距离对鸟类进行观测。这种方法虽然避免了对鸟类的干扰,但需要投入大量的人力物力,还需要观察者有相当的生态学、分类学知识。
通过深度神经网络,AI 可以进行高效的图像识别和声音识别,为鸟类观测提供了新方法。在鸟类主要活动地部署音视频记录设备,设备可以将记录到的数据上传到服务器,随后通过 AI 对这些数据进行分析,提取出音视频中的信息,最终得到鸟类在这一区域的分布。这一方法已被国家林业和草原局广泛应用于公园、湿地和生态保护区中。

图 9:部署在黄河三角洲的鸟类智慧监测系统
同时,AI 的这一技能还可以减轻科研人员的工作负担。AI 可以排除背景和噪音的干扰,专注于图像的特征,迅速解决生态学者难以做出判断的问题。**比如下图中的照片,如果没有任何鸟类知识,很难从纷繁的羽毛中迅速判断出雏鸟的数量。

图 10:一窝雏鸟的照片,你能分辨出图中有多少只雏鸟吗
AI 正广泛应用于鸟类活动监测和鸟类分布分析中,自下而上搭建起鸟类研究的全系统,实现特定区域的「鸟口普查」。相信在 AI 的帮助下,我们能够对生态系统有更透彻的认识,制定出更加符合当地情况的生态政策,逐渐恢复地球的生物多样性,保护我们的地球家园。


 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国