

海洋的氧气水平变化是一个重要的环境指标,受多种因素的影响而发生变化。氧气的溶解和循环受到温度、生物活动、物理环流和化学过程等因素的影响。“海洋与湿地”(OceanWetlands)小编注意到一项新的研究,该研究发现海洋中的氧气含量与气候变化密切相关,可能对海洋生物多样性产生重大影响。这项研究由美国新泽西州普林斯顿大学、巴拿马安孔巴尔博亚史密森热带研究所的科学家们的科学家们共同完成,于2024年初发表在《海洋科学年度评论》的第16卷上。
研究指出,海洋中的氧气含量对地球生命的多样化起到了关键作用。然而,随着人类工业活动的不断发展,氧气水平正面临着巨大的挑战。工业化导致了生物量的过度提取和栖息地的破坏,从而影响了全球能量和营养平衡,进而改变了海洋的氧气平衡。
此外,研究还发现,人类活动引入的营养物质以及全球变暖对海洋氧气消耗产生了直接影响。沿海地区尤其受到影响,尽管它们仅占海洋面积的一小部分,但却维持了大量的海洋生物生存。来自农业活动的营养物质加剧了这些地区的氧气消耗,导致了缺氧条件和大规模死亡事件的发生。
面对这一挑战,科学家呼吁采取综合措施来减缓氧气消耗带来的影响。他们指出,理解气候变化与海洋氧气水平之间的联系对未来的预测以及对海洋生物生活的影响至关重要。要想有效解决这一问题,需要综合考虑实证观察、生理学研究等多方面因素,共同努力保护海洋生态环境的健康和稳定。
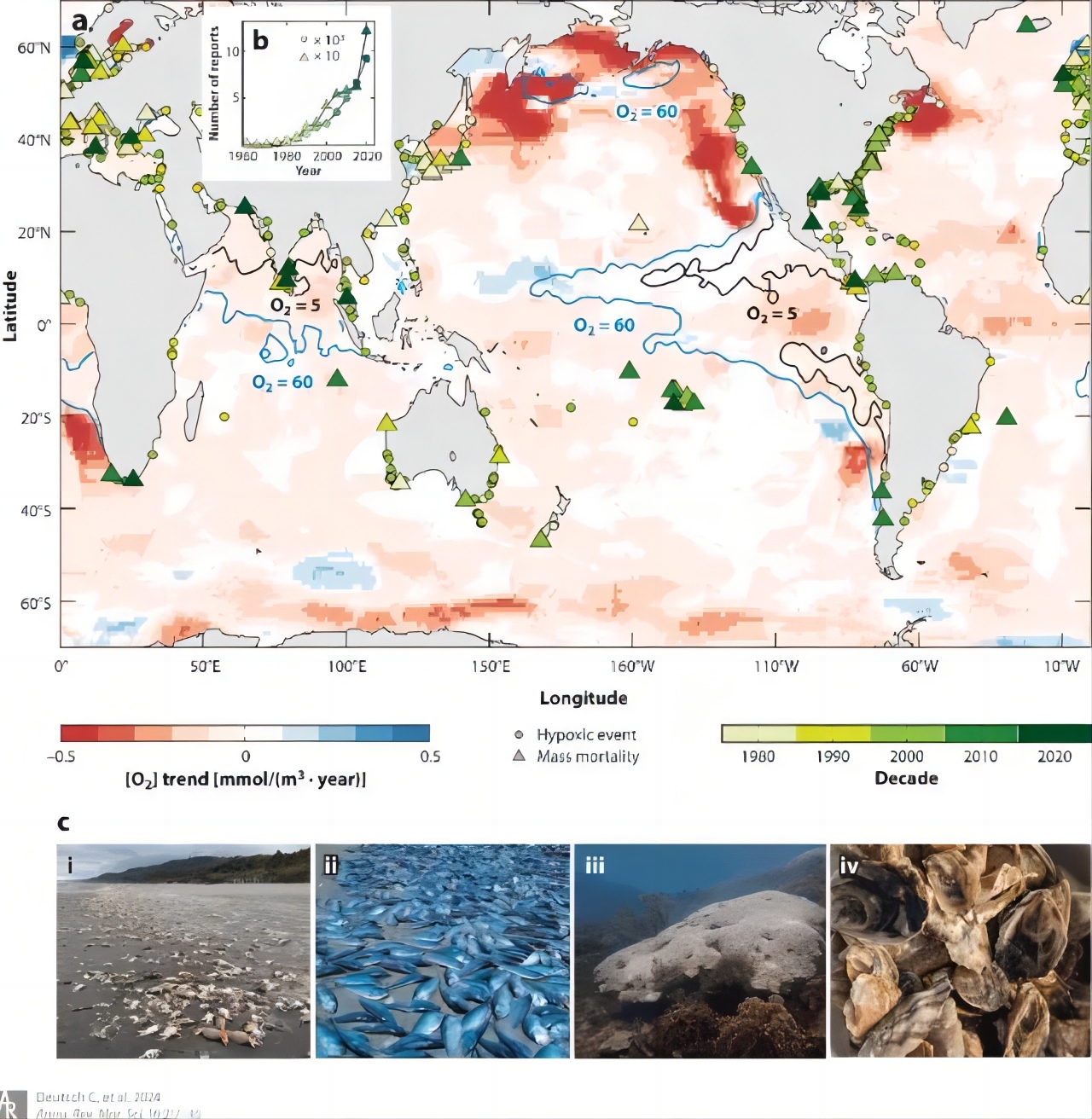
上图:全球氧气浓度([O2])的趋势和极端缺氧事件的发生。在1958年至2015年期间,记录了沿海缺氧事件的历史趋势,这些事件对海洋动物造成了不利影响或导致大规模死亡。彩色区域代表了200米深度处的[O2]趋势,同时叠加了缺氧事件的数据点,其中圆圈表示对海洋动物有害的事件,而三角形表示导致大规模死亡的事件。这些数据来自于Ito等人(2017年)的研究,缺氧事件的数据更新自Diaz等人(2011年)和Altieri等人(2017年)。等高线显示了年均[O2]在上部200米的最小值,分别为5 μM(黑色)和60 μM(蓝色),这些值被普遍认为是微生物和动物有氧功能的阈值。我们观察到,沿海水域的缺氧事件和大规模死亡事件呈加速趋势,每一百次缺氧事件中就有一次会导致大规模死亡。照片展示了受氧气驱动的死亡影响的海洋动物的多样性,包括来自美国太平洋西北海岸的达氏蟹、来自美国罗得岛海岸的油鱼、来自巴拿马加勒比海岸的白化脑珊瑚以及死亡的东方牡蛎。图片来源:Deutsch C, Penn J L, Lucey N. Climate, Oxygen, and the Future of Marine Biodiversity[J]. Annual Review of Marine Science, 2024, 16: 217-245.
海洋的氧气:从全球到微观尺度
在现代海洋中,氧气的含量跨越了地球历史上存在的所有范围,从远古海洋中的缺氧、到海平面上略高于大气压的氧气压力。海洋中氧气的浓度随着纬度从赤道到极地的增加而增加,主要是因为海水中氧气的溶解度会随着温度的降低而增加。在海洋深层(约50米以下),氧气的含量会受到地区水团形成和上升区域的影响,从接近饱和的地区到低值的地区不等。
大部分海洋区域的氧气含量远高于大多数动物的致命水平。然而,这并不意味着海洋中处处都是安全的。缺氧和亚缺氧区域虽然占海洋总体积的极小比例,但却是生物学上极其重要的环境。这些地区几乎不适合大多数海洋动物居住,甚至对大多数需氧细菌也不友好。但是,它们却是一些厌氧微生物的主要栖息地,这些微生物能够将生物可利用的氮转化为惰性氮气。这种过程对全球营养物库的调控和全球初级生产的限制至关重要,尤其是在低纬度地区。
在不同尺度上,海洋中氧气供需之间的平衡关系都非常重要。这种平衡关系对于理解海洋氧气分布、随气候变化的变化以及海洋生物对这些变化的响应至关重要。
近几十年来的历史观测显示,由于人为气候变暖导致的热量被海洋吸收,海洋中的氧气量减少了约2%。这种氧气流失与温度呈强烈关联。在海洋表面,氧气随温度变化的斜率与溶解度的变化趋势相同,导致表面氧气压几乎没有明显变化,但是在深层,这种趋势变得更为陡峭,超出了溶解度单独解释的范围。全球范围内,深层水域中氧气的减少趋势以及氧气/温度关系的斜率也因海洋区域的不同而异。因此,海洋生物在历史上每经历一度的变暖,其氧气损失量可能因其生活的海域深度范围和地理位置而有所不同。
虽然在观测中已经明显地检测到了全球范围内的氧气损失,但是对其潜在原因的归因以及对未来(和过去)气候状态的预测主要依赖于地球系统模型。这些模型能够参数化氧气的混合、产生和消耗过程。尽管这些模型成功地重现了氧气的全球分布和历史变化的一些特征,但也存在着一些系统性的区域性和时间性偏差,尤其是在低氧区域。地球系统模型普遍预测,在全球范围内,氧气供应和需求的速率都会下降,尽管两者都表现出明显的区域性差异。海洋内部通风速率的降低是由于表面变暖和淡化导致的密度层结增加。在全球范围内,表层海洋中的净初级生产力也会下降,尤其是在低纬度地区,这里的氧气供应更为有限。这种营养物供应的减少是由于贸易风的减弱、赤道上升流的变弱以及来自高纬度源水的营养物流失的结合。海洋表面有机物生产速度的减慢导致了深海中微生物呼吸速率的全球性下降。在全球范围内,循环供应的减少超过了呼吸需求的减少,导致了氧气的下降和全球平均氧气压的降低。
地球系统模型的一个普遍预测是,在热带低氧区域,氧气在气候变暖下仍将保持相对稳定,甚至有所增加。这种现象是由于微生物氧气消耗的减少所致。氧气消耗速率的减少会随着气候变暖而积累,在热带温层达到最大值,与气体溶解度的影响相当,因此在很大程度上抵消了氧气供需之间的影响。热带温层中的氧气供应和需求之间的平衡在现代和过去的气候变暖中都有所发现,这可能是由于营养物循环和氧气循环之间的基本耦合。在养分有限的海洋中,表面营养物通量的减少与微生物需求之间存在内在的联系,因为同一物理循环会向表面初级生产者提供养分。
这些机制的组合,导致了热带地区和高纬度地区氧气压趋势的相反。在高纬度地区,由于更完整的表面营养物消耗,导致额外的氧气消耗,使氧气压显著下降超出了气体溶解度的影响范围。相反,在热带地区,氧气饱和度和AOU之间的平衡导致氧气几乎没有变化,在不同的模型中,氧气压变化的趋势并不稳定。然而,对太平洋最大的次缺氧区的重建变化显示出与热带贸易风的密切关系,这些贸易风被预测在气候变暖下减弱,因此未来的次缺氧区应该会缩小,即使全球氧气下降,这一趋势得到了最近地球系统模型分析的确认。在所有模型中,即使是那些预测轻微热带氧气下降的模型,生物吸收的驱动力——氧气压——最终也会上升。这些海洋动态的净效应是氧气从富含氧气的高纬度地区重新分配到低氧热带地区,这对海洋生物有着重要的影响。在全球氧气持续下降的同时,热带地区缺氧的减少意味着厌氧微生物的栖息地可能会减少,从而减弱了水域中营养物的去除作用。与此同时,热带地区海洋动物对垂直迁徙的障碍(即低氧区)可能会减弱,从而部分抵消了较高温度对生物需求的影响。
水生生物的氧气供需平衡
水生生物的氧气供给和需求平衡受到温度的强烈影响。温度影响着生物对氧气的代谢需求和供应的生物物理过程。通过实验发现,动物的氧气消耗速率会随着氧气浓度的降低而减慢,形成一个临界氧气阈值。当氧气浓度低于这个阈值时,动物的代谢活动会受到限制。此外,温度升高会导致动物的氧气需求增加更快,而氧气供应的能力增加较慢。因此,温度越高,动物所需的氧气浓度阈值就越高。
不同物种的氧气耐受性随着温度的变化呈现出广泛的差异。大多数物种的氧气耐受性随温度升高而增加,但增加的速度比代谢率增加的速度要慢。此外,动物的体型大小也可能改变其氧气供需比。体型较小的物种通常对缺氧的耐受性更好。所以说,动物的氧气供需平衡受到温度、氧气浓度和体型的影响,这些因素决定了动物在不同环境条件下的生存和适应能力。
海洋生物的生存与氧气供给有密切关系,受温度影响比较大。一些物种对氧气的需求随温度而变化,这影响了它们在不同水域的生存能力。
温度敏感性较高的物种,通常只能在较冷的水域生存;而温度敏感性较低或者为负的物种,则更适合生活在热带表层水域。此外,物种的活跃缺氧耐受性,也影响了它们的栖息地选择。对于某些物种来说,具有较高的活跃缺氧耐受性,意味着它们能够在低氧水域中生存;而具有较低耐受性的物种,则只能在高氧水域中生存。
海洋物种的地理分布也受到氧气供应的影响。物种的分布范围通常与氧气供应与温度的关系密切相关。温度较高、氧气供应较低的热带水域往往物种较少,而温度较低、氧气供应较高的亚热带水域则物种较多。
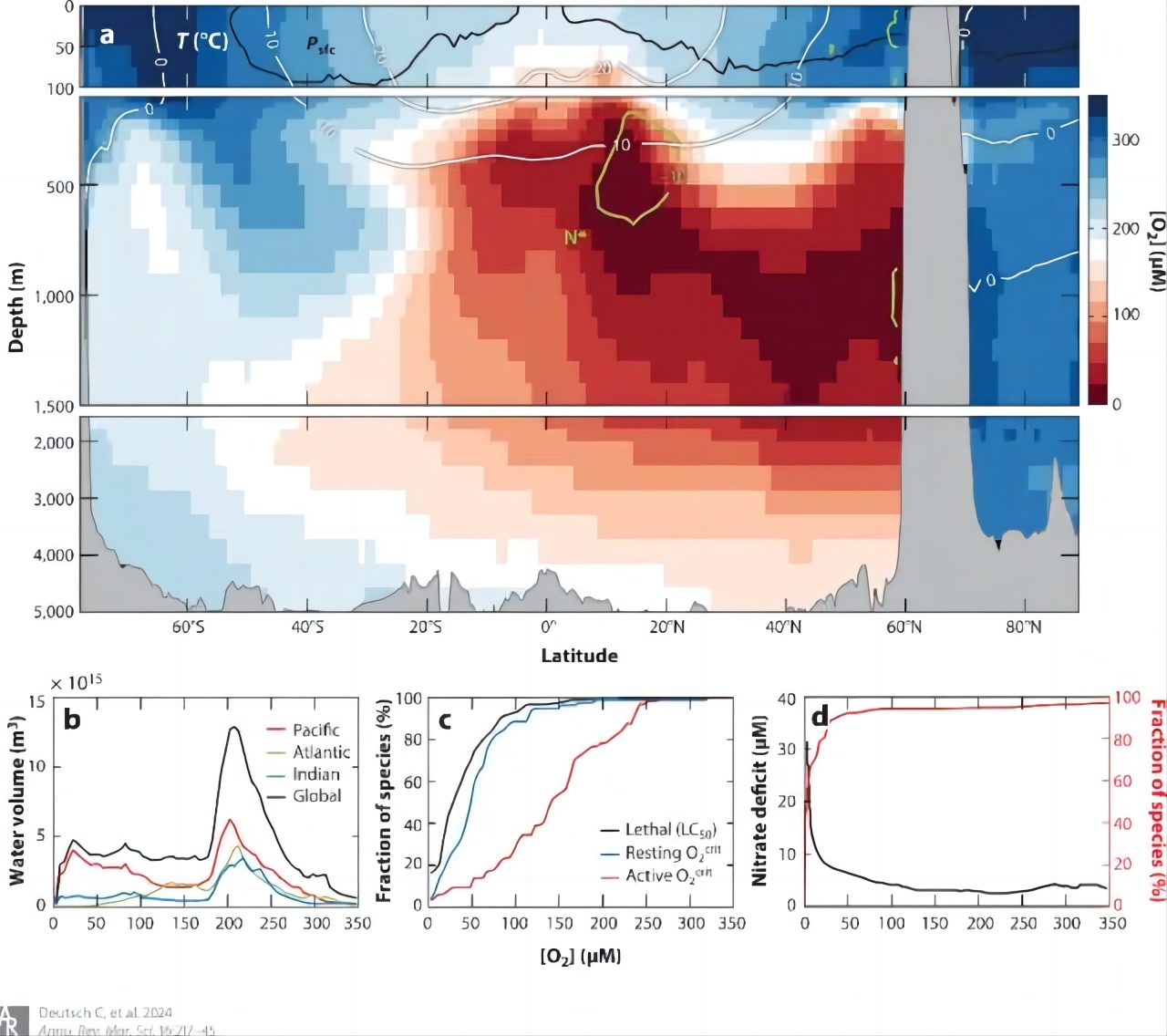
上图:氧气在海洋中的分布和生物对氧气的需求情况。(a) 观察到在东太平洋的不同深度和纬度处,氧气浓度(用颜色表示)随着温度(白色线条)的变化而变化。叠加的绿色等值线显示了氮盐浓度相对于磷酸盐浓度的差异,这是由微生物的氮损失引起的。黑色线条表示氧气与大气之间的平衡位置。(b) 全球海洋中不同氧气浓度下的水体积分布。(c) 累计了动物对氧气需求的数据,包括致死浓度和基本代谢需求。(d) 观察到海洋中氮盐与氧气浓度之间的关系,以及能够在不同氧气浓度下进行氧气摄取的微生物数量。随着氧气浓度的降低,需氧微生物的数量减少,而厌氧微生物的活动则增加。图片来源:Deutsch C, Penn J L, Lucey N. Climate, Oxygen, and the Future of Marine Biodiversity[J]. Annual Review of Marine Science, 2024, 16: 217-245.
气候变暖对海洋生态系统的影响
气候变化对海洋生态系统产生了深远影响,包括缺氧区的变化、以及海洋动物的体型变小。
该研究指出,随着气候变暖,一些最大、且最令人惊讶的海洋生态系统影响,出现在氧气含量最低的区域。模拟显示,随着热带海洋氧气含量的改善,缺氧区可能会减少,与近年来扩张的预期相反。然而,气候模型预测显示,在未来一个世纪,热带地区的氧气含量可能会增加,而更高纬度地区的氧气损失可能会加剧,尤其是北太平洋,这里的氧气已经低到足以支持厌氧代谢。这种变化是模型倾向于使热带地区氧气含量增加的结果。古海洋观测也支持了这种预测,显示出未来气候变暖可能导致热带地区的氮损失减少。这种情况可能会影响氮循环,进而影响海洋生态系统中的生物多样性。然而,对于这些变化的准确评估受到了多种因素的阻碍,包括对缺氧区结构和氮循环过程的模拟的不足。
气候变暖和氧气损失,也被认为可能导致海洋动物的体型缩小。实验研究表明,较低的氧气含量和较高的温度可能导致动物的体型减小。然而,这一假设还存在一些争议,因为氧气供应和需求的同步变化可能会限制动物体型的适应性。尽管如此,根据模型预测,在未来几十年内,一些海洋动物的体型可能会减小,特别是那些构成海洋食物链基础的小型物种。这种体型缩小可能会对海洋生态系统产生广泛影响,包括可能导致一些物种灭绝的风险。
这些变化,可能会对海洋生物多样性和生态系统功能产生重大影响,需要进一步研究和监测。
未来的展望和方向
气温和氧气对物种分布和多样性以及它们对气候变化的反应影响,最为显著的是在全球范围内。然而,生物体和种群生活在高度多样的环境中,在不同的地方,甚至在浅水区域,条件可能在几米的距离和几天甚至一天内发生变化。大尺度气候动态与生物地理的关系仅能在个体生物在同时面临环境和生态压力的空间和时间尺度上预测其命运,这构成了未来的主要不确定性,也为进一步研究提供了机会。
气候对海洋生态系统的适居性的影响,在陆地上也是如此,部分通过极端条件的频率和强度发挥作用。短暂的极端条件,相当于海洋天气,即使不经常发生,也会对种群产生长期影响,从而影响适居性。空间和时间的生境异质性的特征和后果对观测和模型来说都是一个重大科学挑战。通过地球系统模型,这个挑战可以在很大程度上得到解决,这些模型越来越能够以高空间和时间分辨率模拟关键生物应力。这样的模型表明,极端低氧和高温条件的共存在大部分开放海洋上升。
海洋热浪与低氧的短暂极端条件组成了一个完美的代谢风暴,虽然暂时性的,但却显著地侵蚀了有氧适居性。检测这些事件及其长期后果受到高时空分辨率的水文测量的稀缺性的阻碍,尤其是当与受到这些风暴影响的物种的生理特征和生物地理响应相结合时。
最近,生理特征、生物地理野外调查和高频率水文数据的整合揭示了在一个广受关注的加勒比珊瑚礁中代谢风暴的关键作用。对两种栖息于珊瑚礁的脆弱的星形动物进行的实验室实验得出了适应其温暖的热带珊瑚礁环境的生理耐受度和代谢指数特征。尽管珊瑚礁条件平均来说都在两种物种的耐受限度之内,但耐低氧的物种在变化更大的珊瑚礁部分不存在。两种物种之间的生态位分割源于对具有不同特征的物种在代谢风暴中的差异影响。
珊瑚礁代谢风暴的频率和强度随着温度的升高而增加。通过温度和代谢风暴频率和强度之间的经验关系的外推表明,这个加勒比珊瑚礁将逐渐排斥越来越多的低氧耐受物种,尽管历史上氧分压趋势不存在。温度驱动的低氧极端条件的频率或强度增加可能加速其他热带海洋生态系统的生境丧失,即使平均氧分压保持不变,全球模型对大多数热带地区都做出了这样的预测。在热带生态系统中,气候条件相对稳定,动物可能生活在它们的生存空间边缘,这种情况下,海洋天气的影响可能尤为重要。
评估代谢风暴增加是否是普遍现象将需要更持续的高频率测量氧气和温度之间的协变关系以及驻留物种的温度依赖性低氧耐受性的同时知识。填补这些数据空白对于建立海洋天气事件的极端条件与其长期生物影响之间的机制联系至关重要。
由大规模气候变暖导致的氧气损失将通过富营养化增加,即使在东界区域,那里来自上升的营养物质流量自然很高。大规模气候变化模式在开放海洋中产生氧气异常,最终到达沿岸环境,提供一定程度的可预测性。然而,开放海洋水域穿过大陆架断层的过程是复杂的,氧气异常的保存程度仍然不确定。设计用于预测全球气候变化的模型通常不适合研究海岸过程,如中尺度空气海洋相互作用和底栖-海洋相互作用,这些过程发生在更细小的尺度上。
通过船只和(越来越多的)浮标测量的广泛尺度氧气模式的历史采样,可能难以与主要通过锚定和(越来越多的)滑翔机观测的高频率变化的海岸水域联系起来。例如,在报道发生低氧致死性质量事件的地点,最接近的最大全球数据库(世界海洋数据库)中的最低历史氧气水平通常超过100微摩尔/升。在建模和实证分析中,尺度不匹配,仍然是一个重大障碍,而合成数据集允许将大尺度变化无缝地与低氧的地方表现联系起来,这是迫切需要的。
除了高频率的时空变化外,生态和进化压力也有可能修改由于个体到全球尺度上的氧气供应和需求的不平衡而引起的基于特征的适居性丧失的投影。对于温度和氧气的海洋生物多样性的变化通常假定了多样但不互动的物种之间的固定特征和环境位。虽然这样的模型在再现当前和过去海洋的大规模模式方面取得了成功,但它们代表了有限的生态和进化过程,通过静态而不是动态的生理生态参数。特别是,它们对物种响应如何导致重新构造食物网的级联生态效应以及特征适应如何促进一些长期生物多样性损失的逃逸提供了有限的见解。将生态和进化动态纳入量化特征基础模型当然是一个巨大的挑战,但它可以通过已经可用的观察来指导。生活在慢性低氧中的物种提供了一个窥视特征和生态系统如何适应氧极端的机会。
随着有氧性耐受物种的生物地位赢家和输家的出现,高度耐受的物种将填补较不耐受物种留下的生态位,导致功能性生态系统变化。如果它们生态上与消失或脆弱物种通过捕食-被捕食、互利或其他关键相互作用相联系,那么即使是有氧耐受物种也可能会消失。表现为时间不同步的现象,如捕食者-被捕食资源的不匹配,也可能导致营养异步。模拟由不同氧敏感性引起的生态网络对生态影响的影响将需要针对多个相互作用物种和群落的温度依赖性低氧特征的定向测量。
氧气流失和气候变化相关的环境变化的未来与人类对海洋生态系统的大规模干预同时发生。由于人类捕捞压力导致大型物种数量减少和灭绝风险增加的情况已经得到充分记录,可能对对氧敏感性高的顶级捕食者特别严重。随着氧气导致的消失事件的重复性,更小的动物的增加也可能加剧。超过一半(54%)的大规模死亡事件至少发生了一次,导致从生长缓慢到生长快速物种的转变。尺寸结构的变化也与生态系统简化紧密联系(附录A)。这种联系涉及基础或关键物种的丧失,例如牡蛎和珊瑚,它们为鱼类和其他无脊椎动物提供复杂的结构栖息地,并且在近一半的最近大规模死亡事件中已被识别。
对于加剧的有氧约束的进化响应有能力缓解由人类引起的气候驱动的个体氧气供应和需求的不平衡。然而,这样的过程,通常是缓慢和不可预测的。它们也不太可能防止或扭转对直接依赖海洋为生计和文化的人类所造成的损害。未来生物扰动的程度取决于累积的温室气体排放,但氧气趋势和由此产生的生态系统响应可能会在排放减少或停止后继续显现。海洋生物多样性从过去的大规模灭绝中恢复需要大约3-5百万年,这说明了社会对未来几十年的气候选择的地质持久的影响。现在减少二氧化碳排放以避免全球氧气不平衡对全球海洋生物多样性的影响还不算晚,但每延迟一年采取行动就意味着更多的生态系统受到干扰,更多的物种消失。
思考题 | 举一反三
Q1. 氧气流失和气候变化如何影响海洋生态系统中的物种生存与繁衍?这种影响,又可能会如何在不同海洋区域展现出差异性?
Q2. 在面临氧气流失和温度上升的情况下,海洋生物的生理特征和生态环境之间存在怎样的互动关系?这对于生物多样性和生态系统稳定性意味着什么?
Q3.人类对海洋生态系统的大规模干预,会怎样与氧气流失、以及气候变化相互作用?这对于人类的海洋资源利用和保护策略又有何启示?
END
本文仅代表资讯,不代表平台观点。
欢迎转发(请注明来源)。
编译 | 王芊佳
审核 | Sara
排版 | Sara
参考资料略




 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国