

引言:
在面对高压力社会和高压力工作,家庭时,压力是如何让人患上情感障碍—焦虑,抑郁,双相情感和肥胖的?为何童年心理创伤,原生家庭的负面刺激会影响一生?如何面对一段让你有压力的感情和婚姻?如何走出这些阴影?这篇文章从认知神经科学的角度打开了另一个视角—压力和压力激素对大脑产生的负面影响,它们在某些情况下会造成脑损伤和认知损害,以及如何从专业视角看待走出压力,抑郁等问题。

生命体必须适应环境的改变并使自身保持完整才能生存下去。年幼的有机体必须在本体的改变和发展中完成这一过程。在面对外部环境中的应激源或稳态威胁时,个体通过自身的改变维持完整性。这种改变包括整个身体对时间的反应记忆能量和资源的转移,从而促使当下的生存,这种改变经常以抑制生长和修复为代价。因此,负责适应压力的系统与负责生长发育的系统交织在一起,了解这些系统及功能对我们理解发育过程非常重要。

下丘脑-垂体-肾上腺皮质HPA系统通常称为压力系统。尽管HPA系统主要参与压力反应,但它的影响非常之广泛。在无压力情况下,当这一系统维持内环境基础功能稳定时,其发挥积极的作用是支持生长与发育。在挑战性环境下,HPA系统的活动增强,只是激素和肽类物质释放,这些物质会抑制生长和修复过程,转而帮助有机体维持当下生存所必需的功能来应对压力和挑战。
压力和挑战会引起HPA活动增强。个体面临的应激源有两种,即生理应激源和心理应激源。生理应激源例如蓝光刺激,缺氧,生理性创伤,它们涉及脑干系统,并没有认知加工的需要。心理应激源例如控制约束,恐惧条件作用,或者暴露于新异环境,指责谩骂,规则压迫,心理操控,则需要高级认知加工过程的参与。与生理应激不同的是,心理应激并不涉及对生理稳态的威胁,或许我们可以理解为大脑对心理应激的反应是将其定义为对未来生理威胁的预期。生理应激和心理应激涉及了不同的神经回路,其中心理应激主要涉及边缘系统的回路。
一,HPA系统的神经生物学基础
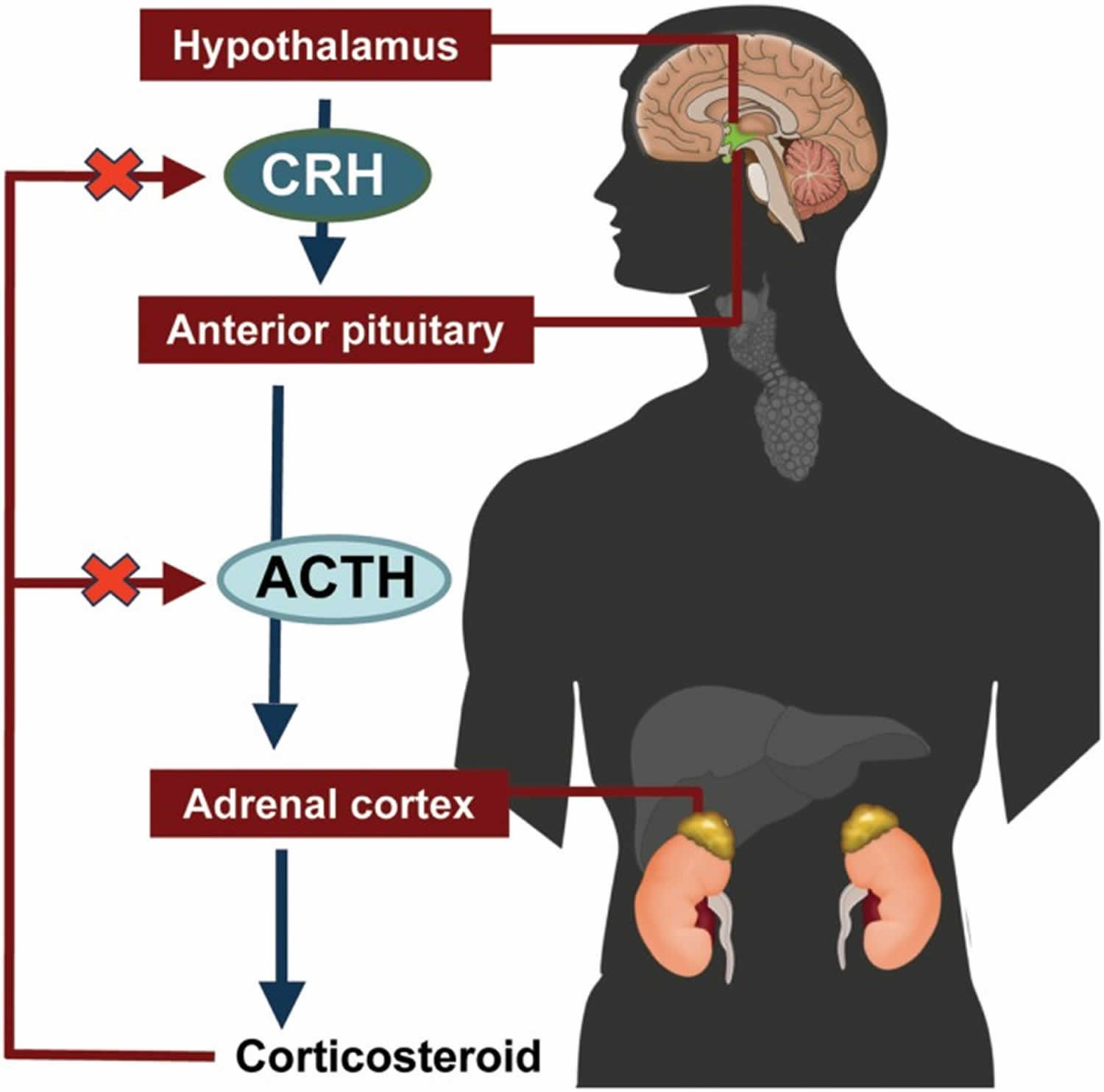
中枢神经系统负责协调内分泌系统面对威胁和挑战时的反应。边缘系统的输入使认知-情绪的挑战能够激活HPA轴。压力和挑战信号首先激活中央下丘脑室旁核,而后刺激CRH(促肾上腺皮质激素释放激素)的释放。CRH和其他促分泌激素例如后叶加压素一起,刺激脑下垂体合成并释放ACTH(促肾上腺皮质激素),而后ACTH通过循环系统到达肾上腺,这时候肾上腺髓质分泌肾上腺素,而肾上腺皮质分泌糖皮质激素。对于人类来说,起主导地位的糖皮质激素是皮质醇。糖皮质激素钝性结合球蛋白,它可以以这种结合方式储存起来,以便随时快速释放。这可能解释了短期休息并不能减压的机理。
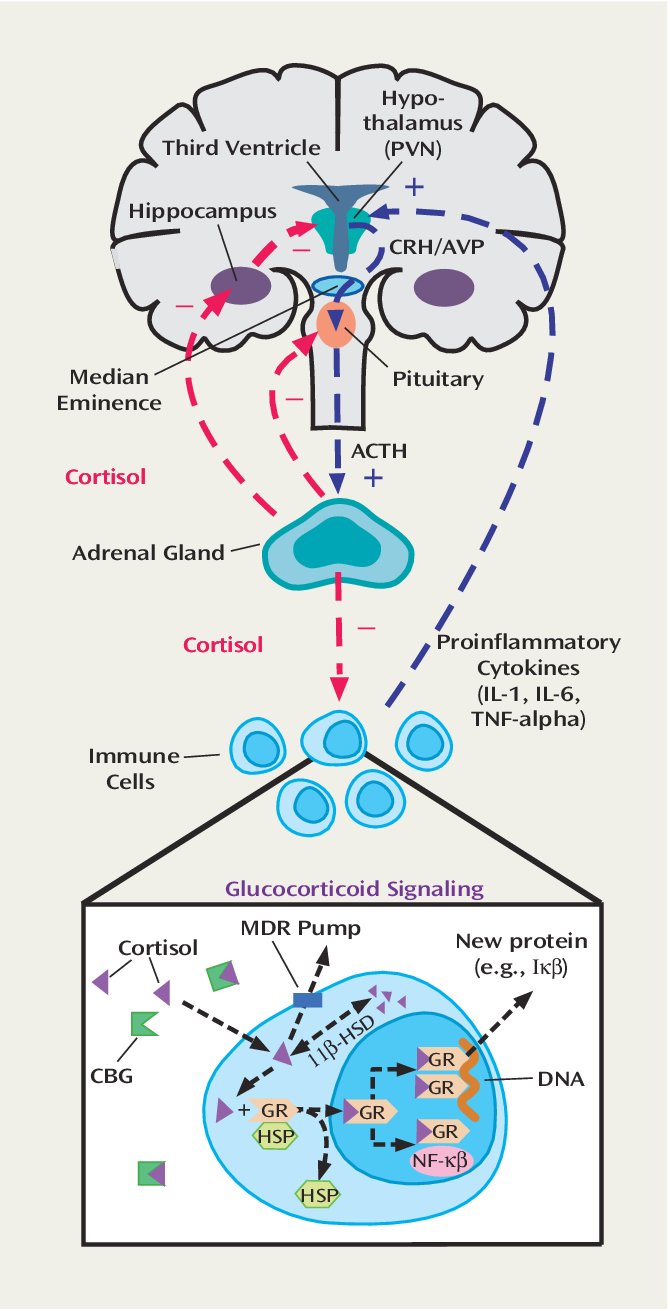
皮质醇影响了几乎所有身体的器官和组织,这种影响包括能量调动和免疫抑制效应。皮质醇可以轻松的通过血脑屏障,而整个中枢神经系统中都遍布皮质醇的受体,换句话说,中枢神经系统对皮质醇尤为敏感,并且从多个方面影响神经系统。
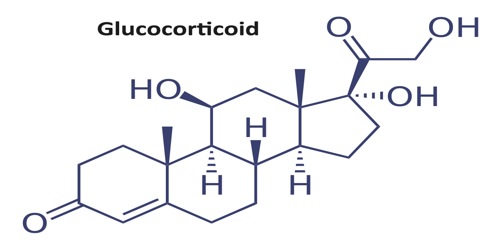
1,HPA系统的活动
HPA系统主要通过边缘系统来对心理应激源产生反应,边缘系统包括杏仁核和杏仁核周边系统。在边缘系统损伤时,面对心理刺激源时抑制了皮质醇的上升,但是这些区域并不影响生理应激源刺激时,例如生病和创伤时皮质醇水平的上升。边缘系统和HPA系统的联系称为LHPA。但尤为重要的是,CRH除了下丘脑它还会在大脑的其他区域产生,杏仁核中具有大量可以CRH的神经元,并且CRH使这一区域糖皮质激素的水平大幅增加,同时与恐惧和焦虑相关的行为也随之增加。这是一个重要提示——压力会导致产生情绪反应,情绪反应又最终促使皮质醇水平增高,增高的皮质醇继而又刺激杏仁核形成正反馈刺激HPA产生皮质醇,因此在后面讲到的负反馈机制完备的情况下,情绪控制对于抑郁和焦虑来说尤为重要,思考是要加载认知负担的,认知负担又会增加压力,因此,对于焦虑和抑郁来说,停止无用的思考和情绪控制对于LHPA调控尤为重要。
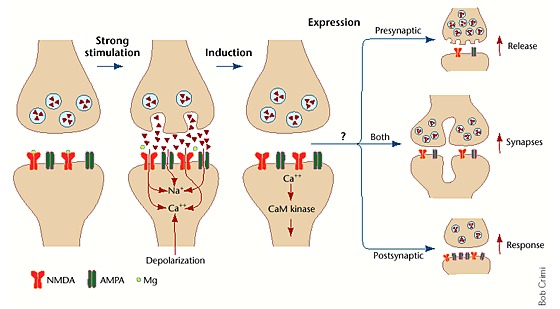
2,糖皮质激素的调控:受体的作用
HPA轴通过负反馈回路调控糖皮质激素水平。糖皮质激素的反馈点位有脑下垂体,下丘脑,海马和额叶皮层。升高的糖皮质激素会抑制下丘脑室旁核CRH的产生。并且对垂体,海马上的两种受体进行压力条件下和基线条件下的负反馈:即盐皮质激素受体,糖皮质激素受体。两者都参与压力反应,并且两者都是必需的,尽管盐皮质激素受体联系更紧密。
但这些受体在人类大脑中的分布仍然是未知的,人类额叶皮层与负反馈的相关程度更高,这种负反馈机制牵制了糖皮质激素面对应激源的反应。
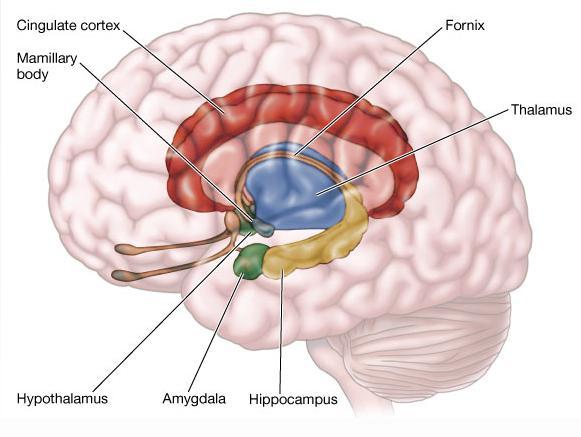
3,LHPA轴的基线活动与LHPA轴的发育
LHPA系统显示出昼夜节律的变化。糖皮质激素产生的基线水平在刚刚醒来时出现峰值,在睡眠开始时达到低估。这种变化与一天开始时需要调动能量有关。
新生儿在面对应激事件时,表现出稳步提升的糖皮质激素水平。但是反应性在2-4个月时明显下降,糖皮质激素反应明显下降的另一个阶段是6-12个月,这个年龄段的婴儿在父母照料时,面对较为温和的应激源不再表现出糖皮质激素水平的升高。

二,早期经验与LHPA轴
脑发育和发展的过程涉及了机体与复杂及具有挑战性环境之间的互动。负责情绪加工的神经生物系统具有可塑性,因此也能够被经验所改变。这些系统在生命早期,边缘系统的连接还在发展时特别容易被塑造。在动物实验中,每天抚摸或亲自隔离的操纵能够永远地改变它们对压力的反应,恐惧行为以及认知功能。

这些早期经验的作用有一部分时通过负反馈过程实现的,这一过程通过糖皮质激素受体数量的变化来调控。研究者发现,母婴交互式LHPA轴发育的重要中介条件。增强母亲关爱程度的操纵使个体成年后应对心理应激源时的反应性降低(表现为降低的糖皮质激素和ATCH水平)。这提示着,儿时得到更好关爱的个体在未来可以表现出更强更灵活的抗压能力。尽管增强母爱关注程度的动物并未表现出相关激素的基线水平差异,但是它们表现出在新异环境中减弱的恐惧反应以及面对多种应激源时降低的LHPA反应,同时伴随糖皮质激素反应的快速终止,这种现象代表着个体具有一种非常有效的负反馈系统。

具有这类经验动物的海马和额叶皮层中具有更多的盐皮质激素受体,糖皮质激素受体。然而打断亲子照顾的操纵具有相反的效应,产生了被称作过度反应的后代,这些后代的糖皮质激素受体水平更低,其LHPA系统调节能力欠佳。

早期剥夺实验也采用了非人类灵长目作为研究对象(最早的剥夺实验是代表行为心理学的华生恐吓实验和母亲剥夺实验)在社会刺激剥夺环境下喂养的幼猴经常表现出增强的恐惧行为。同伴喂养而非母亲喂养的猴子,表现出更多的害羞,支配性低以及对新异性的恐惧,他们面对压力时也会表现出更高的LHPA反应。较轻程度的切断母婴关系也会影响LHPA系统。

探讨人类早期经验与LHPA轴活动关系的研究非常少。在罗马尼亚孤儿院养育的儿童未出现正常儿童身上出现的糖皮质激素昼夜节律。而且,8个月以后被收养的儿童白天出现了更高水平的糖皮质激素,在孤儿院时间越长其糖皮质激素升高的水平越高。
受到多年的虐待后,患有创伤后应激障碍综合症PTSD的儿童也会表现出类似的糖皮质激素水平的长期效应。这些儿童表现出LHPA轴多水平的失调,包括升高的糖皮质激素基线水平。童年期经历过性虐待的成年女性在面对应激源时表现出更大程度的CRH升高。由于研究结果没有报告太多正常经验范围内个体HPA系统的发育情况,因此,这些结果的解释应该谨慎。

在非极端养育条件对LHPA的研究中,检验了自然情况下不同教养方式造成的差异。不安全的依恋关系以不敏感和不稳定的照顾行为为特征,与具有安全依恋关系的儿童相比,具有不安全依恋关系的儿童面对一系列潜在威胁刺激,包括疫苗接种,亲子分离和新异时间时,更可能表现出糖皮质激素水平升高。这
些数据说明了LHPA系统的活动可以受到母亲照顾的影响。但是,儿童自身所扮演的角色也会对安全关系的发展产生影响。

尽管对于人类来说,经验是如何改变后期LHPA轴的活动知之甚少,但是至少存在几种机制,例如边缘系统环路的改变参与调控LHPA,以及盐皮质激素受体和糖皮质激素受体数量的改变。
三,LHPA轴与认知记忆功能损害
1,糖皮质激素对海马神经元损伤的量效关系
海马在多种认知过程中扮演着重要角色,这些认知过程包括学习和记忆。持续的高糖皮质激素水平对海马活动的改变可能导致认知缺陷。
糖皮质激素和海马表现的关系不是线性关系,而呈现倒U型关系,即中等水平的糖皮质激素对海马表现来说是最佳的,而极低和高水平的糖皮质激素会降低海马神经元的活性。
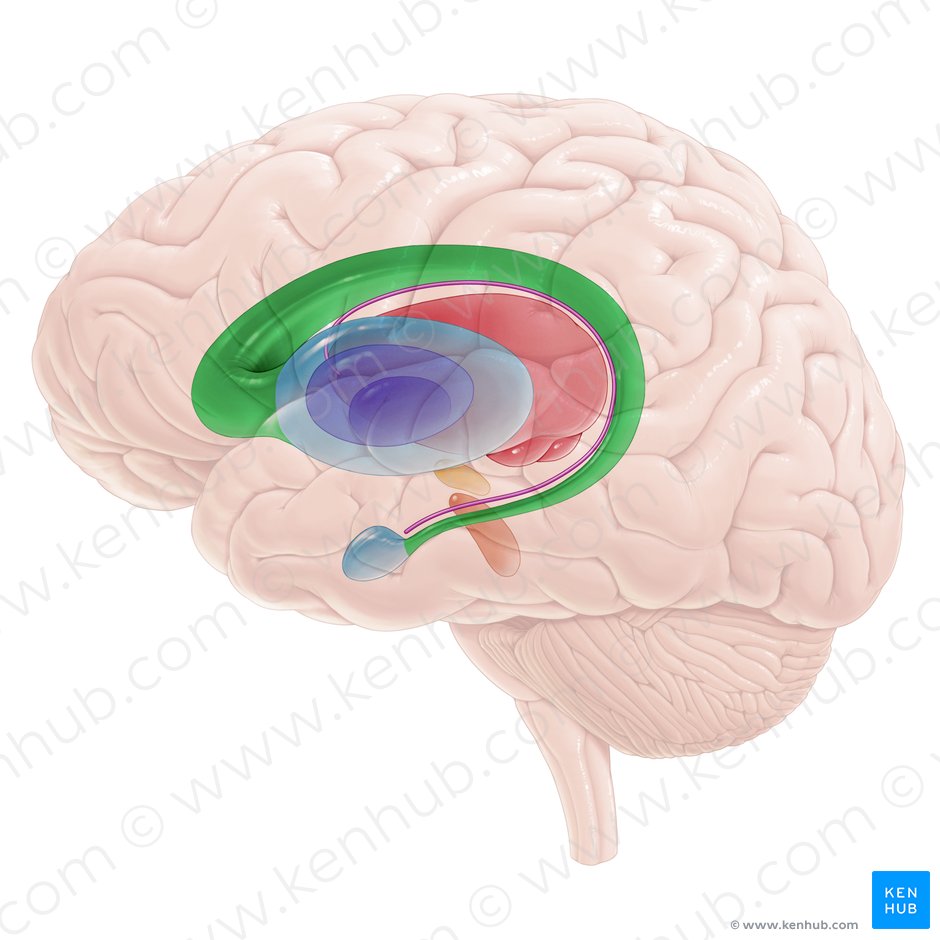
海马神经元的活动调控会影响突触的长时增程。长时增程效应LTP是指持续时间较长的突触效率的提升。这种强化发生接受高频电刺激的时候,这种过程被认为是学习过程所必需的。
证据显示,糖皮质激素以类似的机制对LTP进行调控,糖皮质激素水平极高或者低的情况下会抑制长时增程,而中等水平的糖皮质激素会促进长时增程。这提示着最好的学习状态既不是高压力水平下,也不是极度放松下,而是在中等压力之下。
糖皮质激素不仅仅影响神经活动,有证据表明,持续的和极高的糖皮质激素水平升高会导致海马神经元的退行性改变。此外,慢性的,高水平的糖皮质激素不仅仅损伤神经元,也会影响个体面对其他突发挑战时的生存能力。
证据显示,高水平的兴奋性氨基酸例如谷氨酸盐会激活NMDA受体,这是导致海马突出萎缩的主要元英。而兴奋性氨基酸的释放在一定程度上受到糖皮质激素的调控。慢性压力下糖皮质激素水平升高会产生迁延的谷氨酸盐水平升高,着会导致NMDA受体的持续激活以及潜在的细胞死亡。

糖皮质激素抑制脑中葡萄糖的传输,导致能量缺乏从而使海马难以持续正常的活动过程。这种能量的缺乏可能是糖皮质激素引起海马损伤的一个主要机制。由慢性糖皮质激素水平升高引起的海马萎缩与记忆功能缺陷有一定关联,这种剂量与反应的倒U型曲线时有盐皮质激素和糖皮质激素受体被占据量改变所造成的,因为盐皮质激素受体的绑定提高海马兴奋性,而糖皮质激素受体则相反。
糖皮质激素可能损伤人类的认知功能,急性施用糖皮质激素与依赖于海马的记忆系统损伤有关联。此外类似心理生理压力水平的外源性糖皮质激素施用会抑制成人语言陈述性记忆的功能。这些数据提示,人类记忆表现的损伤可以发生在急性的或持续一段时间的压力下,这种压力可以是从轻到重的不同程度压力。

2,糖皮质激素对前额叶皮层的损伤
证据表明,糖皮质激素与额叶皮层的执行功能有关。升高的糖皮质激素水平可能影响与注意相关的神经系统。升高的糖皮质激素水平与额叶皮层功能障碍有关,例如注意缺陷,而且干扰了诸如计划,注意调控以及工作记忆等额叶功能。
动物实验表明,糖皮质激素水平的慢性增高可能会导致额叶皮层神经的退化变性,高水平的糖皮质激素导致细胞外高水平的兴奋性氨基酸会造成潜在伤害。
尽管证据表明糖皮质激素的增高会影响额叶皮层的功能,但是也可能是前额叶皮层的个体差异影响了LHPA系统的活动。例如,压力造成计划困难,而计划困难继而激活LHPA系统形成压力循环式增加。
很多检测糖皮质激素对认知影响的研究都涉及对成人急性施用糖皮质激素,但对于儿童来说,早期经历的逆境对于认知的影响在幼年时期可能很微小,但是很小的差别也会使儿童进入特殊的发展轨道,使其在后期表现出显著的功能障碍。

四,LHPA轴与焦虑恐惧情绪的正反馈
情绪研究者一直把糖皮质激素作为个体差异的负面情绪(例如恐惧和焦虑)标记物。由于LHPA轴活动与恐惧系统存在关联,杏仁核本身也会在情绪状态下分泌CRH并且聚集高水平的糖皮质激素,因此面对威胁事件时,糖皮质激素升高最明显的个体会表现出最强烈的惊恐或焦虑反应。
但出现了很多相反的研究结果,存在了是否可以将糖皮质激素的活动与负性情绪直接关联的疑问。将糖皮质激素的升高视为一种适应不良的观点是不妥的。糖皮质激素与参与情感加工的神经系统之间存在动态关系。

杏仁核与LHPA轴
面对心理应激源时的LHPA轴活动受到了边缘系统的调控,LHPA也调控边缘系统的情绪加工过程。杏仁核被认为是恐惧诱导的感觉,自动化输入以及行为输出的中枢脑结构。杏仁核中的CRH在增强LHPA轴活动以及中介杏仁核对恐惧的反应中起到了重要作用。

研究发现,提高糖皮质激素的负性生活事件也能够影响下丘脑和杏仁核的CRH水平。杏仁核产生CRH的细胞体中已经发现了糖皮质激素受体(正反馈的)。其中CRH1受体是糖皮质激素的主要受体,它调节焦虑的产生。CRH1受体在猴脑中分布广泛,特别是在边缘系统,而且CRH受体的表达表现出高度可塑性。
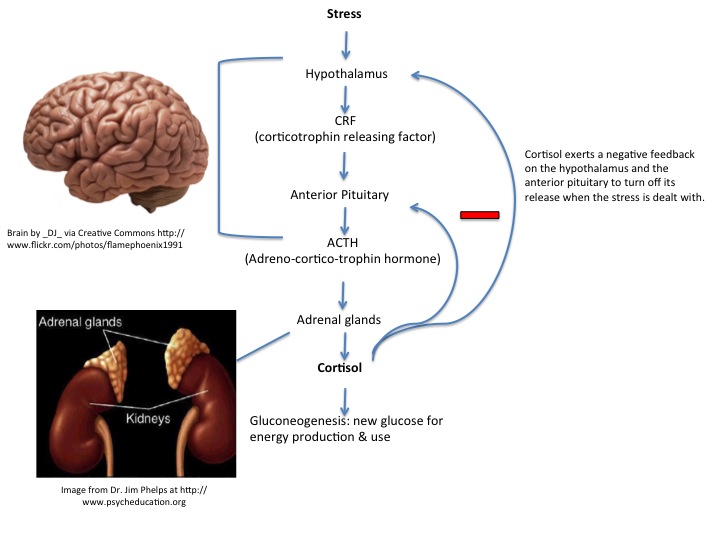
动物实验表明,多种不同的应激源刺激能够导致大鼠脑中CRH1受体的RNA增多。另外剥夺母亲照顾的恒河猴在海马和额叶皮层有更多CRH1受体的表达。这种上行调控可以增强恐惧反应,或者降低恐惧阈值。因此,糖皮质激素和CRH系统的交互作用是一个潜在的机制,压力环境可能通过这一机制来影响个体对未来应激事件的反应性。惊跳反应实验证明了杏仁核中的CRH,循环系统中的糖皮质激素,以及慢性糖皮质激素增高均能增强CRH醋精动物惊跳反射。
这些系统的活动可能影响了病理性焦虑的发展。恐惧,焦虑在一般水平下的个体差异可能与杏仁核或终纹床核的兴奋性有关,差异又可能源于不同水平的CRH。慢性重复出现的压力经历可能影响恐惧回路,并且造成这些回路的长期改变。这些改变随后会降低引发恐惧的刺激阈值。导致潜在的焦虑。PTSD的患者与健康者相比,表现出了更高的CRH基线水平。研究支持了杏仁核核LHPA轴参与增强个体情绪感受敏感性的观点。

五,LHPA轴与脑发展
研究者试图探究发育中个体的LHPA活动与恐惧焦虑的关系,在群体研究中当事件具有威胁性时,糖皮质激素明显升高。这被认为是一种正常的恐惧调节反应,糖皮质激素的升高可能使个体更好的面对恐惧事件。
研究者还考察了个体性格与糖皮质激素之间的关联,有证据表明,显示出逃避,害羞,冻结,惊恐的猴子具有更高的糖皮质激素基线水平;面对威胁和心理挑战时,情绪反应更大的个体也表现出了更高的糖皮质激素水平。焦虑行为与糖皮质激素相关的结果在人类儿童的研究中也有类似发现,儿童糖皮质激素基线水平的升高与行为抑制的增强相关联。

但上面的研究不是绝对成立的,儿童可能会采用保护性策略,即外向行为的个体会通过升高皮质醇来调节新异环境下的行为,而害羞,焦虑的儿童可能发展出应对策略帮助他们进入人群挑战时保持较低的皮质醇水平。
焦虑,抑郁的注意控制可调性
人们对事件的加工方式也会影响糖皮质激素的产生与恐惧及焦虑的关联。例如,人们如果注意到了信息决定了他们对情景的情绪反应和行为反应。能够将注意从威胁性刺激转移并抑制反射性反应的儿童可能成功地运用这种策略来降低他们的焦虑。这种个体差异有时被认为是一种主动的行为控制,代表了一种注意的形式,这种形式使人们能够自主调节自身的行为。

能更好调节注意的个体对引发恐惧,焦虑,抑郁的刺激的敏感性也较低。不同个体中,需要努力控制注意力从不愉快事件中分离出来的个体相比能更好调节注意的个体,表现出了更高的皮质醇水平和更低的刺激阈值。
除了注意调节,环境背景也是中介变量。活跃的个体在新异环境中刚开始表现出了高皮质醇水平,而适应环境后则呈现出较低的糖皮质激素反应性,但是融入不好及出现负性情感的儿童会表现出高水平糖皮质激素反应性。这些证据提示了新异环境刺激对糖皮质激素水平升高相关联,带有威胁性的新异冲突经常与升高的LHPA活动相关联。

尽管重复的暴露与压力环境通常会降低皮质醇的反应,但在有些情景下这种适应并不会发生。当个体在在重复持续的压力环境中,随着时间推移,这些个体在持续面对压力时,不会表现出皮质醇水平下降,相反他们的LHPA系统对这种环境产生了敏感性。而这种改变会提高个体产生发育情感障碍的风险——PTSD,焦虑症,抑郁症,双相情感障碍等。

逆境导致的系统失调会使循环系统中存在着超量的糖皮质激素,这可能是使恐惧,焦虑,抑郁的脑环路发生改变的潜在因素。CRH受体在杏仁核,额叶皮层等处被高水平的皮质醇引发正反馈,继而增高的CRH水平可能导致杏仁核过度敏感,提高了焦虑障碍发生的风险。在具有正常调节能力的系统中,通过负反馈系统,这种高水平的糖皮质激素会正常地降低糖皮质激素的释放至基线水平。但是在失调的系统中,慢性的高水平皮质醇会下行调节参与这种负反馈过程的受体。这可能会损害系统的自动压力关闭能力,从而导致持续的高水平皮质醇激素循环。这一过程的持续存在会造成糖皮质激素相关的病理状态。

对情感障碍的研究者发现,收到过性虐待的女孩表现出低水平的ACTH,但是他们的糖皮质激素基线水平增高。这是因为,脑垂体如果对过量的CRH呈现出低反应,那么系统就会对这种低反应产生代偿——增高CRH和皮质醇。这个研究和成人,儿童抑郁研究结果一致,反馈机制失调。
注:糖皮质激素对代谢的影响:
(1)通过促进糖原异生、减慢葡萄糖分解、减少机体组织对葡萄糖的利用等方面的作用来增高肝糖原,升高血糖;
(2)提高蛋白质的分解代谢,增加血清氨基酸和尿中氮的排泄量,造成负氮平衡。而且大剂量还能抑制蛋白质合成;
(3)可增高血浆胆固醇,激活四肢皮下的酯酶,促使皮下脂肪分解,而重新分布在面部、上胸部、颈背部、腹部和臀部,形成向心性肥胖。
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国