

引文标:李一航.在我们身边的城市生态系统中,物种正在发生怎样的变化?.《生物多样性保护与绿色发展》.百家.2022年6月6日.ISSN2749-9065
摘要:当提到生态系统我们一般想到的会是什么呢?温带阔叶林,寒带泰加林,热带雨林还是沙漠?其实我们自己就身处在生态系统中,只不过是以人类为主的城市生态系统。自然生物为了适应这一生态系统,会根据自身所处的环境选择基因流动,基因漂变,基因突变中的任意一种来改变自己,以便自己能在这样的大环境中得以繁衍生息。疫情期间,如果你一时去不了野外,或者因为平时忙碌而无暇野外,不妨就在城市中探索这独特的生态系统,感受这身边的自然。
关键词:城市生态系统,物种,基因流动,基因漂变,基因突变
城市生态系统定义/特征,以及和演化的关系:城市生态系统,就同字面意思,是由城市为主体的生态系统。不同于任何其它生态系统,城市生态系统几乎完全是人类活动的产物,而且相比较其他生态系统出现的时间要晚很多(因为人类建立城邦的时间从地球历史角度来看并不长),可以说是最年轻的生态系统类型。
同时,城市生态系统也有着许多独有的特征。
首先,该生态系统中有许多不适合生物生存的地方,尤其是各种人造物,如马路表面,玻璃表面等;其次,植被的覆盖率相比于大部分生态系统都较低。相比与自然环境,城市的污染程度明显较高,且有着不同方面的干扰。再者,由于人类活动和有意饲养,非本地物种往往会比自然环境中更多,而其中可能有破坏性大的入侵物种。以上四点都导致城市生态系统的生物多样性是较低的,因为很多生物无法适应这种被高度人为改造的环境。
然而反直觉的是,城市生态系统同时又被认为是促使演化最快的生态系统类型(Johnson & MunshiSouth, 2017)。这不仅是因为当今世界城市的快速发展和扩张,其独有的特征也使得不同生物有了和自然环境截然不同的演化模式和方向,而城市物种的演化又会促使生态系统的改变,从而影响到我们人类的生活。
所以,对于常驻城市的大部分人来说,了解城市物种的演化是至关重要的。这不仅能帮助我们更好地规划城市、控制害虫,同时也有助于达成人与其他生物和谐共生的生态格局。
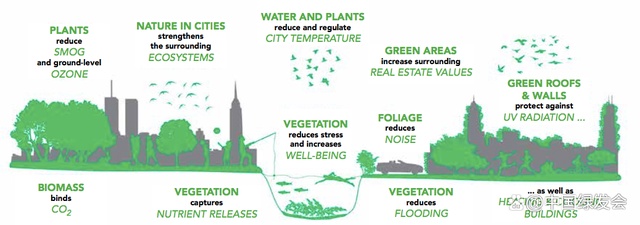
城市生态系统的组成以及生态功能的概述
城市物种的演化主要分为两个方面:非适应性演化和适应性演化
非适应性演化:不涉及自然选择的演化,主要包括基因漂变、基因流动和基因突变。
基因突变(mutation):突变的概念,相信大多数读者都不陌生,这里就不过多赘述。关于城市是否会促使物种突变还没有明确的说法,但许多证据都表明城市的一些特征,如高污染程度和人为干扰,可能会增加突变的概率。
拿最极端的情况举例:福岛和切尔诺贝利地区的生物都有着很高的突变概率。此外,城市物种的突变原因,可能并非由城市本身的环境所导致。比如一些自然状态下存在的突变,可能因为适应城市环境而稳定存留下来。最经典的例子,莫过于英国工业革命时期的桦尺蠖:黑色的个体比白色个体数量更多。是因为树皮被工业废料染成了黑色,使得这些个体拥有了很好的保护色,从而能更有效躲避天敌攻击。这个案例通常被拿来解释自然选择,也就是适应性进化的一部分,但黑色个体其实并非完全是自然选择的结果,而是之前就已经存在的突变,只不过工业革命创造出的环境更适合拥有这种突变的个体生存,从而使该特征成为了主流。
目前,仍然缺乏对城市环境和突变之间的联系的研究,因此还不好下任何定论,但如果城市确实增加了突变几率,我们就得仔细权衡利弊了。因为有些自然环境中不受偏好的突变,可能会在城市环境保存下来,而其存在,可能对自身种群或者其他物种有不利影响。再比如某些害虫的突变在城市中大规模扩散,我们的健康也会受影响。

两种类型的桦尺蠖Biston betularia,黑色个体更加适应工业革命时期,灰色个体更适应工业革命以外的时期。
基因漂变(genetic drift):指的是特定基因在种群中比例的改变,主要有开拓者效应(founder effect)和瓶颈效应(bottleneck effect)。前者是指由带有亲代群体中部分等位基因的少数个体来到一个全新环境并重新建立新的群体,使得该群体中所有个体之间的基因差异较小、但整个群体与其原栖息地其它群体间差异较大。一些物种从城市周围的环境迁徙至城市后,会在城市中定居并繁衍后代,久而久之,城市种群就与其他自然种群有了比较大的基因差异。
同样,同一种群在不同时期进入城市,都有可能独立触发开拓者效应,而城市生态系统中的各种人工屏障和不适宜的地区都会导致栖息地碎片化,使得物种之间的基因交流变得更加困难,进一步加大种群之间的基因差异并缩小种群内的基因差异。
不同于开拓者效应,瓶颈效应则是指一个完整的种群经历了某些变化后大幅缩小,使得只有少数个体存活,而使得那些由少数个体组建的新的种群也是彼此之间基因差异小,且和不同的种群基因差异大。在城市生态系统中,瓶颈效应一般是由杀虫剂等针对性措施所导致的。不论是害虫还是其它受影响的生物,都会有相当一部分被杀虫剂消灭掉,而由那些幸存个体组成的新群体,就很容易触发瓶颈效应。
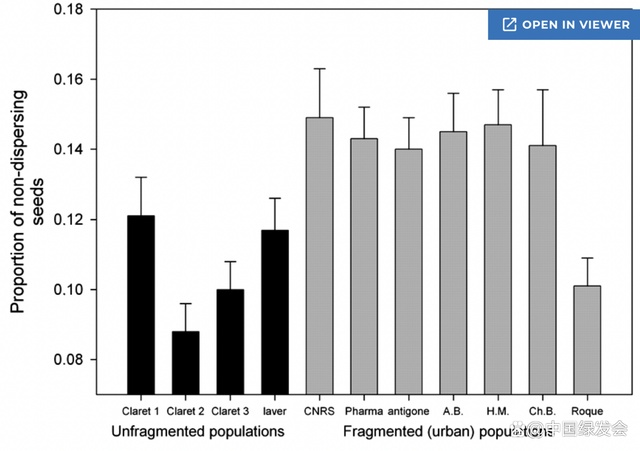
关于城市物种的基因漂变有不少研究,但仍然有一些需要解决的问题。其中一个便是原生物种是否比外来物种更容易经历基因漂变。
基因流动(genetic flow): 指的是不同种群之间的基因交流。正常的基因流动,能让种群内的个体之间保持较高的基因多样性,而种群与种群之间的基因差异相对较小。正如前文所说,城市生态系统让不同种群之间的交流更加困难,所以基因流动也会减少。对于那些扩散能力差的物种尤其是如此,两个种群可能数十代都不会有任何交流。基因交流对于种群的健康至关重要,所以交流频率低的种群一般会更加脆弱。为了增加基因交流,修建生态通道和公园等设施是很好的解决方案。
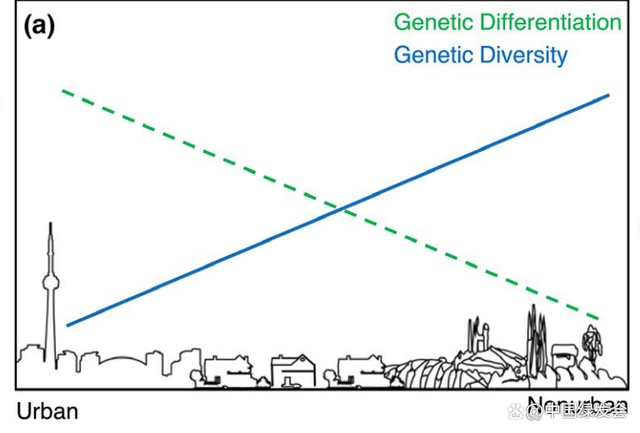
主张城市生态系统会导致物种基因交流减少、基因漂变增加的假说,被称为Urban Fragmentation model, 在167篇有关城市物种基因的研究中有大于60%的支持了该假说(Miles, Rivkin, et al., 2019)。
值得注意的是,城市物种的基因交流和漂变,并不都是一致的,在城市生态系统中有些物种可能与其他物种有着截然不同的模式。那些与人类有密切联系的外来物种,比如褐家鼠等鼠类,在城市生态系统中反而有更多的基因交流机会。它们一般不会被马路等障碍所阻挡,而人类的交通工具又会进一步帮助其种群进行扩散交流。而其它和人类有长期联系的物种,比如人类驯养的动物以及身上的寄生虫等,也会在城市中有更多扩散和基因交流的机会。这类模式被称为Urban Facilitation model.

相比自然更喜欢城市环境的褐家鼠(Rattus norvegicus)、人类驯化的岩鸽(Columba livia)、以及以类血液为食的臭虫(Cimex lectularius)在城市地区都有着更高的基因交流,算是城市适生物种。
不只是外来物种,一些原生物种也会因为城市化而增强基因交流以及减少基因漂变。
研究者(Miles, Dyer, et al., 2018)对美国西部三个城市的西部寡妇蜘蛛(Latrodectus hesperus)调查后发现:拉斯维加斯和凤凰城之间的种群基因相似度较高,甚至比这些城市种群和城市周围的自然种群之间的差异要低不少。这种蜘蛛在自然条件下只喜欢特定的环境,比如岩石下和干旱河谷附近,而环境的零碎分布使得不同种群之间的基因交流十分有限,促使了漂变的发生。相比之下,城市生态系统有更多理想的栖息地,比如墙角和缝隙间,而交通工具和货物运输又增加了城市间种群的基因交流,这才导致了该结果。
当然,城市之间的差异也是重要的干扰因素。研究中来自新墨西哥州城市阿尔帕克基的蜘蛛就与拉斯维加斯和凤凰城的个体有不小的基因差异,反而更加接近非城市地区的种群。
再次,由于城市和非城市地区蜘蛛在基因上的显著差异,它们的行为模式也有着一定差异。研究者们观察到城市里的蜘蛛更喜欢聚集,而且对同类和猎物有着更强的攻击倾向。如西部寡妇蜘蛛,以它们的毒液对哺乳类效果显著,被归为一类卫生害虫。其对城市环境的强大适应力以及种群间的交流,使得它们并不容易被控制,而长期生活在城市导致的其行为上的改变,又可能加剧它们对于人类的威胁。在未来,人们应该根据其非适应性演化趋势发掘更有效的控制方法。

西部寡妇蜘蛛广泛分布于美国西部,对人类有一定威胁。我之前晚上在洛杉矶的一个市中心公园看到好多这种蜘蛛沿着墙织网,可见其对城市环境的适应……
适应性演化:这类演化模式,相信是大部分读者所熟悉的。因为一般初高中甚至大学的生物课都是拿适应性演化举例。最经典的莫过于达尔文提出的自然选择理论。
自然选择:城市生态系统的很多特征,比如不适合物种生存的表面以及和自然环境不同的温度、气候等,都会对物种施加较强的选择压力,从而促使自然选择发生。
拿植物来举例,研究显示:城市地区的还阳参(Crepis sancta)种群,相比于自然环境的种群,更倾向于产生没有扩散能力的种子。这可能是因为城市的许多建筑会阻挡种子的扩散,或者种子会落在不能生长的地方,如马路上等。在这种选择压力下,产生不扩散种子的个体,将会受到自然选择的偏爱。因为这不仅能增加后代的生存几率,同时还能节省能量——因为一般来说,有扩散能力的种子会需要更多能量来生成帮助扩散的结构(想想看有伞和没伞的蒲公英)。
很多时候,自然选择在动物身上体现的更加明显。在波多黎各的城市中,安乐蜥(Anolis cristatellus)普遍有着更长的四肢和爪垫,有助于攀爬光滑的表面,尤其是各种人造物。城市环境对自然选择的促进是毋庸置疑的,但有时候也很难判断某些特征是自然选择的结果还是非适应性演化的结果。
此外,城市的哪些因素导致了自然选择,选择的特征是只有几个特定的还是在不同条件下有很多不同....这些问题都需要大量的观察和实验来回答。


1)安乐蜥,一种在美洲广布的常见蜥蜴2)还阳参,类似路边小野花的感觉3)在城市和非城市地区还阳参非扩散种子的统计,可以明显看出城市地区的种群产出更多非扩散种子(Cheptou, Carrue, Rouifed, & Cantarel, 2008)。
趋同/平行演化:虽然城市之间,因为地理、文化、政治等因素会有一些差异,但整体来看城市与城市之间的差异是小于城市与周围自然环境的差异的,因此不同地区的城市物种可能会展现出趋同的演化模式来适应相似的环境。
一个比较经典的案例,是不同城市的白三叶草(Trifolium repens),都一定程度丢失了生氰作用(cyanogenesis),而这种作用主要是为了防御草食动物,同时也会降低其抗寒性。城市环境中的草食动物相对较少,但冬天温度普遍更低,因此丢失了这种作用的三叶草将会在城市中更有优势,该基因也会迅速扩散开来。
此外,不同城市的加拿大底鳉(Fundulus heteroclitus),也都不止一次演化出了对污水和毒素的抗性,而这显然是为了适应城市的水质而导致的。还有一些更容易观察的现象,比如很多城市的鸟类和哺乳动物对人类的警戒心都不太强,可能是由于长期与人类生活的结果,当然这是基于大多数人类不会特意伤害它们的前提。(我们学校的松鼠就很不怕人,有些甚至贴脸了也不逃走,更有甚者甚至能从人手里接食。)这些行为上的改变,很多时候都有基因改变做为基础,而基因改变恰恰是自然选择发生的力证。
研究者在欧洲12个城市和相应的非城市地区调查了乌鸫的基因组,发现城市地区的个体在SERT基因上和非城市个体有着显著差异,而这个基因主要被认为与警戒性、危险回避、攻击性等有联系( Mueller et al., 2013)。其中10个城市的乌鸫(Turdus merula)的该基因表达都低于非城市地区,说明城市环境的自然选择,偏好那些拥有更低戒备的乌鸫。
最后,我们可以根据城市之间的相似性做出两种猜测:1,同一物种在不同城市会朝着相近方向演化。2,不同物种在同一城市会朝着相近方向演化,尤其是栖息地和习性相似的物种。关于前者已经有很多例子可以佐证,但后者似乎还缺乏相关研究,读者朋友们将来要是有兴趣也可以观察研究一下。
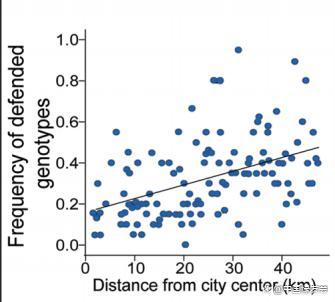

1)白三叶草的防御基因表型与城市中心距离的联系,可以看出离城市越近,该基因表型越罕见(Thompson et al., 2016,)2)白三叶草。3)加拿大底鳉 4)乌鸫
趋异演化:与趋同演化不同,城市内的物种也可能会朝着不同方向演化。
设想一下繁华街道、中心公园、垃圾场和废弃的荒地这4种环境,是不是觉得它们之间的差异并不小呢。城市内部的环境差异,往往比我们想的大,而城市物种为了能在这些环境更好生存,就会各自产生区域性的适应。
区域性适应的产生原因,还是要归功于上文提到的栖息地碎片化:人为(如铁路、马路)或自然(如河流,湖泊)的障碍阻止了物种之间的扩散和基因交流,久而久之,对其所在环境的适应会愈发加强并固定,而与其他环境的同一物种之间的差别也会愈发变大。
这一点,有点类似于地理隔离导致的物种分化,但城市中人造物导致的隔离,一般比自然的隔离更加普遍。关于城市物种趋异演化的研究并不多,主要是因为区域适应性的观察难度和界定标准,以及不好判断区域性适应是否为自然选择还是表型可塑性,亦或者基因漂变所产生的。总之,这方面研究在未来还有很大的空间。

一项关于豚草( Ambrosia artemisiifolia)的研究显示:城市种群不仅和郊区种群在基因和生命周期上有着显著差异,城市种群内部之间也有着不小差异,而这可能是由于不同环境所导致的区域性适应(Gorton et al., 2018)。
物种形成:在我看来,物种的形成是生态演化中最有魅力的一部分,就像宝可梦到了一定时机会“进化”一样。虽然人们对物种形成的机制和速率的研究还并不充分,但已经有不少证据表明城市生态系统在其中有着推波助澜的作用。有些学者甚至认为城市中物种分化、形成的速度,在所有生态系统中是最快的。
那么,是什么因素推动了物种演化呢?
首先是生态分化。同一物种为了适应不同环境而演化出地域性适应(上文提过),久而久之彼此之间在习性、栖息地等方面产生了显著差异以及生殖隔离,也就没办法产生后代了。研究表明亚利桑那州城市地区的家朱雀(Carpodacus mexicanus),演化出了更适应在城市觅食的鸟喙,而其副产物就是求偶歌声的变化(Badyaev, A.V. et al,2008),这使得它们和原先的种群“失去了共同语言”,彼此间的交流也会愈发减少。如果时间足够长,新的雀类物种可能会产生。此外,有一些比较常见的城市物种行为改变,也会对生态分化产生影响,比如更倾向于在城市定居而不是迁徙。

其次是由突变和基因漂变引起的物种分化,属于非适应性演化的范围。
突变很好理解:因为某些因素使一部分种群和其他种群有了区别,彼此之间不再能产生后代,从而诞生新物种。
基因漂变,主要体现在瓶颈效应和开拓者效应上。城市地区的物种容易经历瓶颈效应,而它们所具有的基因型在原先种群中可能只是非常小众的一种。又因为栖息地碎片化等隔离,那些具有罕见基因型的种群将会从原先种群中慢慢孤立出去,最后演变为新物种。
一个比较有趣的案例,是研究者发现:各自独立适应城市环境、生活在三个不同城市的穴鸮(Athene cunicularia),开拓者效应使这三座城市的鸮与非城市地区的种群差异较大,而城市彼此间差异较小(Mueller, J.C. et al,2018)。然而,针对各自城市的地方性适应,可能也会使不同城市之间的鸮基因差异变大,前提是它们能在城市中存活足够长时间。
一个更加有说服力的案例,来自于伦敦的地下库蚊(Culex pipiens molestus)。这种蚊子被认为是由尖音库蚊(Culex pipiens)分化而来的,而其分化的原因是伦敦地下铁的修建。大约150年前,一小部分尖音库蚊选择在地下铁隧道定居,因此和地面的种群产生了隔离,而目前两者已经很难杂交了,即使杂交成功产下的后代也基本都是不孕不育的(Thompson, Rieseberg, & Schluter, 2018)。生殖隔离的产生,是物种分化最典型的特征之一,而这可能只需要短短(在演化尺度上)150年。尽管它现在还只是尖音库蚊的亚种,但只要时间尺度够长且两者间没有交流,分化成独立物种是完全可能的。
有趣的是,地下库蚊对与地面完全不同的环境演化出了专门的适应器。尖音库蚊主要以鸟类血液为食,但地下几乎没有任何鸟类,因此地下库蚊选择以哺乳类为食(老鼠、做地铁的人)。由于地下食物相对缺乏,它们演化出了不吸血就能产卵的能力。同时,地下相对稳定的温度,使冬天不会太寒冷,它们也就退化掉了滞育能力(类似于冬眠)。地下库蚊虽然老家在伦敦,但现在已经扩散到世界很多地方了。其在新的栖息地是否可以和尖音库蚊杂交还有待研究。

1)尖音库蚊,你曾经打死的某只蚊子可能就是这种 2)地下库蚊,你去国外旅游坐地铁时可能打死过
最后,我认为最有意思的一种,即杂交产生的新物种。
城市生态系统因为人类活动密集,有不少不属于本地生态的外来物种会被强行带入到本地生态系统中,而其中有一些可能亲缘关系和本地物种比较近,也就是说可以产生有繁育能力的后代。
对,就是自然状态下完全不可能碰面的两个物种因为人类搭桥,而得以见面并产出许多后代……这可不是什么浪漫故事,而是很可怕的事情。
那些杂交而成的后代,都会多少排挤它们的父母,使得两个物种在同一个栖息地数量下降,甚至完全被这个新的“缝合怪”所取代。有些超级后代甚至会变成入侵物种,对其他生态系统产生破坏。
这可不是什么臆想,而是有确实案例支撑的。来自欧洲的两种杜父鱼(Cottus rhenanus和Cottus perifretum)所杂交产生的后代,侵占了大部分原本不属于它们父母生存范围的水域(Renaut, 2011),对当地物种造成了一定的影响,而杂交的产生又与水利工程等人为因素有一定关联。虽然这起案例并没有发生在城市,但城市物种也能通过相同的原理,即人为干涉,导致其与本不该接触的近亲物种杂交,产生出可能具有潜在危害的后代。



1)Cottus perifretum 大斑杜父鱼2)cottus rhenanus莱茵河杜父鱼3)两者的杂交后代


 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国