前庭器官的“星际迷航”——生物系统如何计算惯性运动新华网-科技前沿大师谈 2016-05-03 作者:刘丫 |
【星际迷航】
科幻影视系列星际迷航的柯克船长带领船员乘坐企业者号飞船穿梭在浩瀚的宇宙,寻找新的世界、发现新的文明,勇敢地前往前人未至之地。试想有一天,企业者号被卷入一片未知的空间,飞船的导航系统处于瘫痪状态,且被切断了与外界任何形式的信息交流,船员们被迫启用手动操作模式。为了减少能量消耗,船员们需要实时监控飞船的运动状态,从而合理调度引擎的功率。因为没有任何外界参照,他们只能利用飞船内部物体的相对运动关系来确定飞船的运动信息。经过几个小实验,他们惊喜地发现要推动的积木的个数越多,难度越大(见图一),这意味着惯性定律在这个空间依然是适用的。惯性度量的是一个物体抵抗外界干扰,保持自己当前状态(静止或者匀速直线运动)不变的趋势。在他们的小实验中,积木的个数越多,惯性越大,因此推动它们难度也就更大一些。
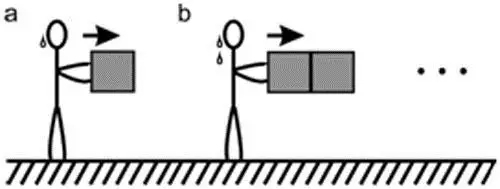
图一,推动物体的难度正比于物体的惯性,惯性的大小用质量表征。(a)推动一块积木可能需要消费一滴“汗”;(b)推动两块块积木可能需要消费两滴“汗”。飞船中的积木处于失重状态,船员通过魔术胶带使自己的双脚粘在飞船的底部。
于是,柯克船长和史巴克找来两块积木,非常仔细地将它俩叠在一起,以保证这两块积木的初始运动状态是一样的(图二 a ),其中“下”被粘在飞船底部,而“上”没有固定在飞船的任何部件,可以“随心所欲”依从自己的惯性。

图二,利用飞船内物体的惯性导致的相对运动来判断飞船的运动状况,箭头表示企业者号的船头。
等待飞船运动一段时间后,柯克等人可能观察到积木的三种叠加结果,通过上下积木的错位情况判断飞船的运动状态。如果他们观察到“上”与“下”并驾齐驱,他们可能要非常恼火了,因为利用惯性原理,飞船内部的人是无法区分静止和匀速运动这两种情况的。
类似的,企业者号的船员们可以毫不含糊地判断飞船是否正在转动:由于惯性作用,不受外力的物体(“上”)将保持自己的状态,从而相对飞船的局部(“下”)沿切线方向飞出去。
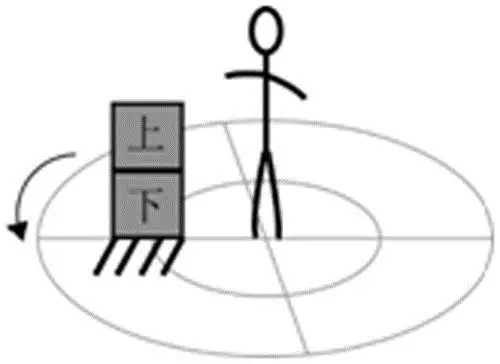
图三,利用飞船内物体的惯性导致的相对运动来判断飞船是否旋转。
判断自己所在系统的运动状态,对于迷失在未知空间的企业者号船员是生死攸关的事情,对于定居在拥挤的地球的形形色色的动物来说,也是不可忽视的小事。
【迷失在内耳的前庭器官】
在芳草萋萋的非洲大草原,正在享用嫩草的羚羊们突然被草丛中的一丝动静惊破了胆,撒开蹄子四散逃跑。一只刚成年的捕猎经验不足的猎豹懊恼地看着四散的羊群,奔向一只看起来跑不动的羚羊(图四)。在残酷的动物世界,对自身运动和环境中其他动物的运动的把握决定着在命悬一线的时刻是劫后余生,还是命丧黄泉!那么,动物是如何处理运动信息的呢?
在演化之初,动物们可能不关心能否解析自身运动。演化史上较为原始的动物(如原生动物等)运动能力有限,它们自身就是环境的一部分,在液体环境中过着随波逐流的生活。对于运动的感知,他们不需要区分敌我,只需知道“敌动推我动”即可。

图四,羚羊与猎豹。一步之遥,将决定羚羊是命丧“豹”口,还是绝处逢生。
随着动物越来越复杂,他们和环境的差别越来越大,能够实现的运动形式也越来越多。这时“敌动推我动”的信念将不再适用,动物们必须想办法区分自身相对环境的运动。此外,演化的四大驱动力(打斗、逃跑、捕食、繁殖)也迫使动物分清自己和环境中其他动物的运动状态,从而采取相应的行为。动物可以利用多种感觉信息输入(视觉、听觉和前庭觉)来检测运动信息,其中前庭器官就是利用惯性原理来检测自身运动的。前庭器官在演化史上约5亿年前出现,随着动物由水生走向陆生,这一器官又衍生出听觉器官。
前庭器官被包埋于颞骨深处(图五),如同迷失在外太空的企业者号。这里(内耳)暗无天日,地形错综复杂,被称为前庭迷路。骨质迷路的内部紧贴着一层膜状结构,膜的两侧流淌着组分不同的内淋巴液。内淋巴液中丰富的电解质离子( K+ , Na+ )使将自身惯性运动的变化转变为电信号成为可能。
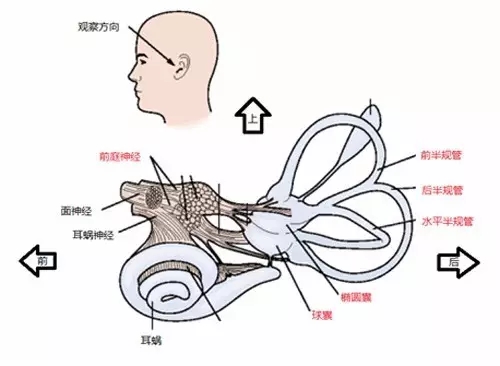
图五,前庭器官的解剖结构。改编自 Kandel, E. R., et al. (2000)。
前庭器官(单侧)包括五个组成部分:两个膨大的囊状结构(球囊、椭圆囊)和三个相互垂直的半圆形的管状结构(半规管)。其中球囊和椭圆囊负责检测线性加速度,半规管负责检测旋转加速度,它们将检测到的信号转化为电信号,经由前庭传入神经上行到脑干等处进行进一步的加工处理。
那么,前庭器官是如何将惯性运动信号转化成电信号的呢?
【前庭器官编码惯性运动加速度】
进一步放大前庭器官的解剖结构,我们将发现前庭器官的感觉上皮上布满了呈六角形结构的纤毛束(图六)。一簇纤毛束对着一个毛细胞,而毛细胞与前庭传入神经存在突触连接。在一个毛细胞中,纤毛依据纤毛的长短呈阶梯状排列,其中最长的一根为动纤毛,其余的为静纤毛。
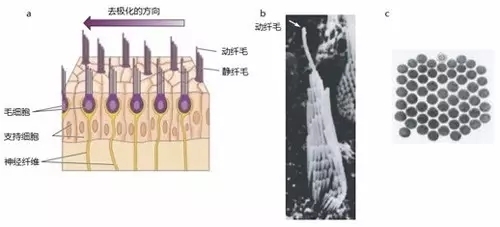
图六,毛细胞的解剖结构。(a)前庭器官的一个切面,在这个切面中所有毛细胞的动纤毛都在同一侧。(b)豚鼠前庭毛细胞的纤毛束的结构。从当前视角可以看到在从静纤毛到动纤毛(箭头所指)的过程中,纤毛的长度逐渐变长。(c)纤毛束的横切面。改编自 PURVES, D., et al. (2004)。
纤毛束的这种不对称排列使得毛细胞具有极性,静纤毛倒向动纤毛使得毛细胞发生去极化(即毛细胞内外的电位差变小),毛细胞释放的神经递质增加,前庭传入神经发放脉冲的频率增加(图七)。类似的,静纤毛倒离动纤毛将降低前庭传入神经发放脉冲的频率。发放频率改变的程度取决于纤毛束的偏折程度。因此,毛细胞可以将纤毛束的偏折方向和程度转化为前庭传入神经发放脉冲频率的改变。
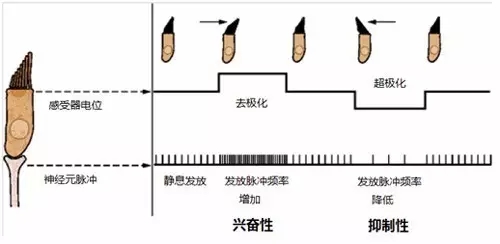
图七,毛细胞将纤毛束的机械偏折转化为前庭传入神经电活动的变化。改编自 Kandel, E. R., et al. (2000)。
前庭器官中的球囊、椭圆囊负责检测线性加速度,它们的感觉上皮的毛细胞的纤毛束所处的基质上布满了微小的碳酸钙晶体(图八 a ),因此球囊和椭圆囊也合称耳石器官。相对于毛细胞外的果冻状基质,耳石的密度要大一些,惯性也要大一些。当头部突然加速时(如向右加速,图八 b ),耳石惯性大、运动滞后,将引起毛细胞偏折(向左)。毛细胞的偏折方向取决于线加速方向,偏折程度取决于线加速度大小。这就是线加速度信息如何转化为前庭传入神经的发放脉冲改变的。从整个群体出发,耳石器官中的毛细胞的极化方向非常复杂,形成的极化向量遍布整个三维空间的任何方向。由于生活在地表的动物持续地受重力加速度的影响,不管我们处于什么样的运动状态,总有部分耳石传入神经处于兴奋/抑制状态。根据爱因斯坦的加速度重力等效原理,耳石器官也可以编码重力加速度,这就是我们能轻而易举区分自己身体倾斜程度的原因。
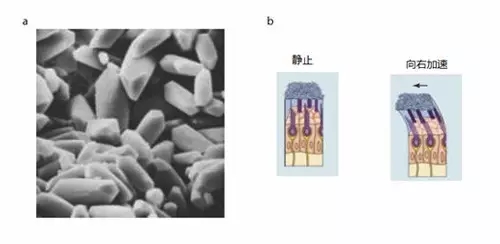
图八,耳石器官传导线加速度。(a)猫的椭圆囊中的碳酸钙晶体的扫描电镜图。每个晶体约50 um 。(b)线性加速运动将引起毛细胞偏折向加速度的方向。改编自 PURVES, D., et al. (2004)。
半规管同样利用惯性原理来检测旋转加速度。半规管充满了内淋巴液,它的基部膨大,被称为壶腹。半规管的感觉上皮位于壶腹,被称为壶腹脊。毛细胞的纤毛束伸入一团凝胶状的结构,也就是壶腹帽。壶腹帽像一个密封不太好的橡皮活塞一样横在壶腹,可以在一定程度上阻碍内淋巴液的流动(图九 a )。
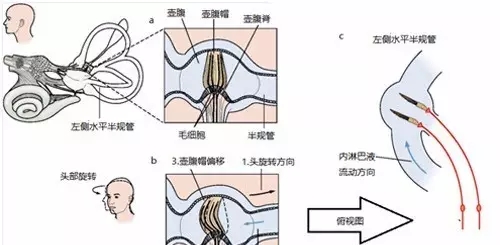
图九,半规管传导旋转加速度。(a)水平半规管在静息时壶腹帽的状态。(b)水平半规管在头旋转时壶腹帽的状态。(c)水平半规管在头旋转时壶腹帽的状态,俯视图。改编自 Kandel, E. R., et al. (2000)。
当头部突然旋转时,内淋巴液由于惯性将流向反方向,进而挤压到挡在壶腹的壶腹帽,引起壶腹帽发生形变,伸入壶腹帽的毛细胞纤毛束也发生偏折,实现旋转加速度向中枢的传导(图九 b , c )。当动物只进行线性运动时,壶腹帽两侧的内淋巴液流向相反,壶腹帽受力平衡,不会引起毛细胞纤维束发生偏折,因此半规管无法传导动物自身的线性运动。不同于耳石毛细胞多样性的极化方向,每个半规管的壶腹中的毛细胞中的极化方向是一样的。通过组合三对半规管传入神经的发放水平,半规管系统能编码三维空间中任意一个旋转方向。
如同企业者号的船员无法区分静止和匀速运动,前庭器官面对这个问题同样束手无策。但我们在日常生活中却很少有这样的困惑,诚然一系列的研究表明,随着惯性运动信息从外周到中枢上行,前庭系统通过整合来自耳石器官和半规管的信息逐渐倾向于编码惯性运动的速度和位置信息(图十)。除了整合前庭系统内的信息,前庭中枢也可以整合来自视觉等其他模态的信息帮助动物精准地判断自身惯性运动信息。
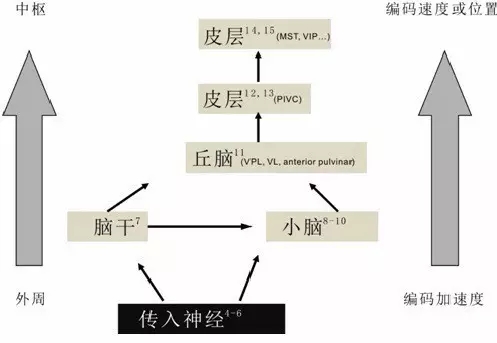
图十,前庭中枢编码惯性运动的速度和位置信息。
更多精彩!欢迎关注“科普中国-科技前沿大师谈”官方微信(kjqydst)。

责任编辑:linzhennan
 科普中国APP
科普中国APP
 科普中国微信
科普中国微信
 科普中国微博
科普中国微博






最新文章
-
为何太阳系所有行星都在同一平面上旋转?
新浪科技 2021-09-29
-
我国学者揭示早期宇宙星际间重元素起源之谜
中国科学报 2021-09-29
-
比“胖五”更能扛!我国新一代载人运载火箭要来了
科技日报 2021-09-29
-
5G演进已开始,6G研究正进行
光明日报 2021-09-28
-
“早期暗能量”或让宇宙年轻10亿岁
科技日报 2021-09-28
-
5G、大数据、人工智能,看看现代交通的创新元素
新华网 2021-09-28





微信扫一扫:分享
微信里点“发现”,扫一下
二维码便可将本文分享至朋友圈。