

蛋白质形成过程中的翻译终止
当核糖体通过移位读到终止密码子时,蛋白质合成进入终止阶段,由释放因子协助终止翻译。
(一)终止过程
当核糖体沿着mRNA分子移动到A位出现终止密码子时,没有相应的氨酰tRNA与之对应,一种释放因子(Release Factor,RF)与终止密码子及核糖体A位结合,另一种释放因子随之结合,改变肽酰转移酶的特异性,催化P位肽酰tRNA水解,从而使肽链从核糖体上释放,如图1所示。接下来,释放因子促使脱酰tRNA脱离核糖体。核糖体解离成亚基并脱离mRNA。
(二)释放因子
大肠杆菌有3种释放因子(RF):RF1、RF2和RF3。RF1识别终止密码子UAA和UAG。RF2识别终止密码子UAA和UGA,RF3不识别终止密码子,但具有核糖体依赖性GTP酶活性,与GTP结合后可以协助RFI或RF2使翻译终止。
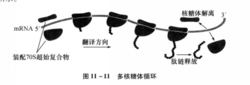
(三)多核糖体循环
当原核生物及真核生物合成蛋白质时,许多核糖体会同时结合在一个mRNA分子上,形成多核糖体结构,进行翻译;此外,核糖体在一轮翻译完成后可以回到mRNA的5’端,重新装配,开始新一轮翻译合成,形成循环。多核糖体循环大大地提高了翻译的效率,如图2所示1。
翻译终止效率蛋白表达水平受许多不同因素和过程影响,蛋白稳定性、mRNA稳定性和翻译效率在蛋白生产和积累中起主要作用。翻译过程分为起始、延伸和终止3个期,对于翻译的起始,原核mRNA需要5'端非翻译前导序列中一段叫Shine—Dalgarno序列的特异核糖体结合序列,在真核细胞中,有效的起始依赖于围绕在起始密码子ATG上下游的一段叫Kozak序列的前导序列。密码子的利用或偏爱对延伸有深刻的影响,如果mRNA有很多成簇的稀有密码子,这可能对核糖体的运动速度造成负面影响,大大降低了蛋白表达水平。翻译终止是蛋白生产必需的一步,但其对蛋白表达水平的影响还没有完全研究清楚,最近的科学研究表明终止对蛋白表达水平有很大的影响,更有效的翻译终止导致更好的蛋白表达。
绝大多数生物都有偏爱的围绕终止密码子的序列框架。酵母和哺乳动物偏爱的终止密码子分别是uAA和UGA,单子叶植物最常利用UGA,而昆虫和大肠杆菌倾向于用UAA。翻译终止效率可能受紧接着终止密码子的下游碱基和紧靠终止密码子的上游序列影响,在酵母中通过改变围绕终止密码子的局部序列框架,翻译终止效率可能被减低几百倍。对于UGA和UAA,紧接着终止密码子的下游碱基对有效终止的影响力大小次序为G>U,A>C;对于UAG是U、A>C>G。
对于大肠杆菌,翻译终止效率可因终止密码子及临近的下游碱基的不同而显著不同,从80%(UAAU)到7%(UGAC)。对于UAAN和UAGN系列,终止密码子下游碱基对翻译的有效终止的影响力大小次序为U>G>A、C。UAG极少被大肠杆菌利用,相比UAAN和UGAN,UAG表现了有效的终止,但其后的碱基对有效终止的影响力为G>U,A>C。对于哺乳动物,偏爱的终止密码子为UGA,其后的碱基可以对体内翻译终止有8倍的影响(A、G>>C、U)。对于UAAN系列,体内终止效率可以有70倍的差别,UGAN系列为8倍。如果终止密码子附近序列没有最佳化,可能发生明显增加的翻译通读,因此减少了蛋白表达。
总的来说,翻译起始框架、翻译终止序列框架和密码子利用应该仔细选择,以利于蛋白的最高水平表达。翻译终止序列框架能几倍地改变蛋白表达水平2。
基因工程中目的基因的定义与结构基因工程主要是通过人工的方法,分离、改造、扩增并表达生物的特定基因,以获得有价值的基因产物。目的基因的分离是基因工程操作的第一步。
通常把插入到载体内的非自身的DNA片段称为“外源基因”(foreign gene)。目的基因(objective gene),又叫靶基因(target gene),是指根据基因工程的目的而设计所需要的某些DNA分子的片段,它含有一种或几种遗传信息的全套密码(code)。
食品原料种类繁多,DNA分子结构复杂,每个DNA分子所包含的基因也很多,要从数以万计的核苷酸序列中挑选出非常小的令人感兴趣的目的基因,是基因工程中的难题。
一般来说,一个具有功能的基因包括基因的启动子区域、编码区和转录终止区。不同类型的生物,其基因的结构有所差别。原核生物的基因组成包括启动子区域、基因编码区域、SD序列区和转录区、翻译终止区。其中SD序列与原核生物基因的翻译有关,基因编码区没有内含子。真核生物的基因包括启动子区域、基因编码区域、翻译终止区,其中基因编码区包括内含子和外显子,蛋白质的序列由外显子的序列决定。因此,在基因工程具体操作过程中,要针对具体来源的基因结构进行分析,进行有针对性的实验。另外,在食品基因工程中,主要是对基因的编码区域进行分离、重组和表达,因此本书所提及的目的基因主要是指基因的编码区域3。

 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国