

群落稳定性的含义
群落的稳定性具有两层含义:群落的抗干扰能力,即抵抗力稳定性;群落受到干扰后恢复到原平衡态的能力,即恢复力稳定性。对于群落的稳定性,有平衡说(equilibrium)和非平衡说(non-equilibrium)两种对立的观点。前者认为,共同生活在同一群落的物种处于一种稳定状态,通过竞争、捕食和互利共生等种间相互作用而形成相互牵制的整体,导致生物群落具有全局稳定性特点;在稳定状态下物种组成和各种群数量都变化不大;群落实际上出现变化是由于环境的变化即干扰所引起。后者认为,组成群落的物种始终处在不断的变化之中,自然界中的群落不存在全局稳定性,有的只是群落的抵抗性(群落抵抗外界干扰的能力)和恢复性(群落在受干扰后恢复到原来状态的能力),其重要依据就是中度干扰理论,即中等程度的干扰水平能维持髙的物种多样性;干扰过于频繁,则先锋种不能发展到演替中期,多样性较低;干扰间隔期过长,演替发展到顶极,多样性又降低; 只有中等程度的干扰使多样性维持最高水平,它允许更多的物种入侵和定居。
群落的多样性是群落稳定性的一个重要尺度。当一个群落有很多物种,而且每个种的个体比较均匀地分布时,物种之间就形成了比较复杂的相互关系。这样群落对于环境的变化或来自群落内部种群的波动,由于有一个较强大的反馈系统,从而得到较大的缓冲。另外,多样性高的群落,食物链和食物网更加趋于复杂,群落内部的能量流动的途径更多一些,如果某一条途径受到干扰堵塞不通时,可能有其他路线予以补偿。因此,多样性髙的群落相对比较稳定。反之,若物种数目少,各个种的相对丰度又不均匀,则群落的多样性就较低,其稳定性也相对较差。
群落多样性与稳定性的关系物种多样性是指地球上生物有机体(生命)的多样化程度,可以分为遗传多样性、物种多样性和生态系统多样性3个层次。对群落而言,物种多样性尤为重要。物种多样性有两种含义:其一是种的数目或丰富度,即一个群落或生境中物种数目的多寡;其二是种的均匀度,即一个群落或生境中全部物种个体数目的分配状况,它反映的是物种个体数目分配的均匀程度。自20世纪50年代以来群落多样性和稳定性的关系一直是生态学界争论的焦点问题,目 前存在两种不同的理论:MacArthur-Elton假说(群落多样性和稳定性正相关)和May假说(群落多样性和稳定性无关)。
MacAnhur-Elton假说的核心是物种较多的群落就可能保持稳定。Mac- Anhur(1955)首次提出了这一观点,认为自然群落的稳定性取决于两个方面的因素:一是物种的多少,二是物种间相互作用的大小,而物种的多少对稳定性的。这个观点可以在物种间的捕食关系中得到实证:当某一物种 数量极多时,必须有大最的捕食者来分散过多的能量,同时又必须有大量被捕食者物种以保证一个物种的数量不会下降过多;当某一物种数量较少时,该物种的每一捕食者必须有大量的替代被食者,以减少压力。Elton根据物种入侵的研究得出了类似的结论,认为一个相对简单的植物或动物群落易于受到毁灭性干扰,因而抵御外来种入侵的能力较弱。
20世纪70年代,MacArthur-Elton假说的生物多样性越高,稳定性越强的观点受到了挑战。May(1973,1976)等认为,随机构造的复杂系统只有在某一临界值内稳定,超过该临界值,系统就不会稳定;在多样化的系统中,一个生态关系复杂的网络,可导致种群急剧波动,而不是更加稳定;共栖的多物种群落,某物种的波动往往会牵连到整个群落;从减少害虫种群暴发的意义上讲,多样性与稳定性之间并无一般的相关关系。他们提出了多样性的产生是由于自然的扰动和演化两者联系的结果,环境的多变性和不可测性使物种产生了繁殖与生活型的多样化。
Pimm(1984)指出,形成多样性-稳定性的两种不同观点的主要原因在于多样性、复杂性和稳定性有许多不同的定义和概念。MacArthur-Elton的理论虽已被许多人加以论证,但还有待于深入和提高;May所提出的理论又过多地依赖于数学方法,而且他所使用的系统又是以人工模拟为主,因而离实际相差甚远。Pimm对多样性-稳定性关系做了一个总结,认为理论上对于一个有更多物种的群落:要使群落更加稳定,就需要物种间的联结变得更少;群落内种群的弹性变得更小;当一个物种丢失以后,群落内种类成分和生物量有较大变化;一个物种丢失以后的状态将保持更长的时间。由此可见,传统的静态的内部自调稳定性概念,难以客观地反映生态系统的动态特征。 Diamond和Case(1986)提出,生态学本身就是一门“多元学科”,在理论和 方法上采用多元论的观点是很必要的。现代生态学已由强调群落或生态系统的平衡、稳定性、均匀性、均质性、确定性、单尺度和小尺度现象,发展到开始重视非平衡、不稳定性、异质性、不确定性、多尺度和等级特点,这将有助于更好地理解多样性-稳定性关系。1
群落稳定性的分析方法食物网与稳定性MacArthur (1955)根据群落的复杂性与群落的稳定性有关的观点,认为一个群落的物种越多或食物网的环节越多,群落内部物种之间的能流途径也就越多,从而群落将更加稳定。于是,他通过一个假想的捕食者与被食者(或害虫与天敌)的营养联系对群落的稳定性作了定量研究。
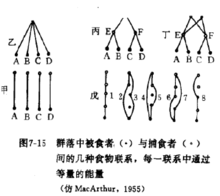
图中甲、乙、丙、丁是四个不同的群落营养结构,其中捕食者与被食者的关系很不一样。每图中A、B、C、D代表被食者,E、F、G、H代表捕食者,每一条线表示捕食联系, 也就是能流途径。甲图中捕食者与被食者的关系十分专一,是单种联系;乙图中捕食者G可以捕食被食者A、B、C、D中的任何一种;丙图是两个营养阶层的捕食关系,G可以捕食 E、F,而E、F又分别可以捕食A、B和C丁图是一个错综复杂的二个营养阶层交错的捕食关系。假设每条食物链能流的量都相同,则甲、乙、丙的食物链都是四条,甲为 AE、BF、CG 和 DH,乙为 AG、BG、CG 和 DG,丙为 AEG、BEG、CFG 和 DFG,每 一条能流的量各占1/4;而丁的食物链有八条,即 AEG、AEH、BEG、BEH、CFG、CFH、 DFG和DFH,每一条能流的量为1/8。
MacArthur的理论和方法,虽然得到不少学者的赞同和支持,如Paine (1966, 1969) 提供的材料。但是,这个方法目 前在实际工作中应用很少,理论上也受到挑战,因为能流在食物网中的分配是不均匀的,难以准确地测定每一条食物链所占有的能量比例。
群落动态与稳定性70年代以来,由于新兴横断学科的发展和不同学科之间的相互渗透,不少学者(如 Angelis,1975; May,1971,1976; Svirezhevetal,1978)基于描述群落动态特性的数学模型,采用系统理论中的Mapunov稳定性定理来分析和探讨生物群落的稳定注。2

 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国