

信号肽信号肽的结构
穿过合成所在的细胞到其他组织细胞去的蛋白质, 统称为分泌性蛋白质( secretory proteins) 。信号肽位于分泌蛋白的N 端, 一般由15~30 个氨基酸组成包括三个区: 一个带正电的N 末端, 称为碱性氨基末端; 一个中间疏水序列, 以中性氨基酸为主, 能够形成一段α螺旋结构, 它是信号肽的主要功能区;一个较长的带负电荷的C 末端, 含小分子氨基酸, 是信号序列切割位点, 也称加工区。当信号肽序列合成后, 被信号识别颗粒( SRP) 所识别, 蛋白质合成暂停或减缓, 信号识别颗粒将核糖体携带至内质网上, 蛋白质合成重新开始。在信号肽的引导下, 新合成的蛋白质进入内质网腔, 而信号肽序列则在信号肽酶的作用下被切除。近年来一些研究表明, 许多分泌蛋白的移位信息虽确由一部分疏水肽段所携带, 但这一部份肽段可以不在N 端, 如终止转运序列存在于新生肽链的C 端, 也可以不被信号肽酶切除, 如卵清蛋白含有内部信号肽, 它的前体与成熟形式都没有被信号肽酶切除的过程, 其N-端氨基酸结构在第9 位有带电基团,疏水结构并不明显1。
信号肽的功能在布洛贝尔“信号假说”的启示和有关蛋白质氨基酸序列的比较研究基础上, 一些新的信号肽先后被发现。这些信号肽负责不同类型蛋白质的新生肽链的定位。信号肽的功能, 已不仅决定一个蛋白质是否为分泌蛋白, 而且和蛋白质或其新生肽链在细胞内的全方位的定位有关。新生肽链或蛋白质中, 一些残基的化学修饰也是转译后加工的一个重要内容。发生修饰的残基决不是任意的, 也和肽链中的氨基酸序列密切相关。从这个意义而言, 这些和残基修饰有关的肽段, 也可认为是残基修饰的信号肽。例如, 在肽链中连接有N-糖链的天冬酰胺残基(N) , 一定是位于Nx( S/T) 这种特定三肽序列中的天冬酰胺; 一些蛋白质的C 末端附近可以为萜类所修饰, 能接上萜类的半胱氨酸酸残基, 同样是具有特定序列的C 末端四肽中的半胱氨酸酸残基,而且其中某些残基还决定了所接上的萜类的长度。为此, 可认为在肽链中尚存在着与残基修饰相关的信号肽。
一些肽段的存在与含该肽段肽链的降解有关, 这类肽段可以视为肽链降解的信号肽。如细胞质中某些带有KFERQ/RIDKQ 序列的蛋白质易于进入溶酶体,然后在那里被降解。又如, 细胞质内快速被降解的蛋白质通常含有PEST 四肽序列1。
原核分泌表达大肠杆菌大肠杆菌是用来表达重组蛋白的首选宿主之一。大肠杆菌遗传背景清楚,操作简单,生长周期短,成本低,易于大规模培养,已经用来表达多种酶制剂和药物制剂,目前市场上有40% 的重组制剂由大肠杆菌表达生产。即便如此,由于大肠杆菌的翻译后修饰能力较弱,在表达多数结构复杂的蛋白过程中会导致蛋白折叠错误聚集形成无活性的包涵体,严重影响蛋白表达量。目前多数蛋白在大肠杆菌胞内表达,需要通过破碎处理获取目的蛋白,且宿主胞内背景蛋白影响蛋白纯化。但利用大肠杆菌分泌系统表达重组蛋白能提高蛋白稳定性及可溶性,保留蛋白N 端真实性,减少宿主代谢负担,有效简化蛋白纯化过程,节省生产成本2。
目前采用的信号肽一般来自大肠杆菌的外膜蛋白( 如外膜蛋白A OmpA 、OmpF、λ噬菌体受体LamB、热稳定肠毒素STⅡ等) 或周质蛋白( 如碱性磷酸酶PhoA、麦芽糖结合蛋白MBP、DsbA 等) , 此外金黄色葡萄球菌蛋白A 和胡萝卜软腐欧文氏果胶酶裂解酶( PelB) 的信号肽也常被采用1。
常用分泌型载体:
pTA1529:大肠杆菌phoA(碱性磷酸酶)启动子和信号肽序列;外源基因插入到信号肽之后;在磷酸盐饥饿时外源基因表达;由周质信号肽酶切下信号肽
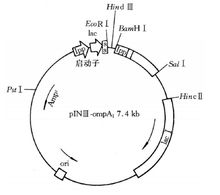 pINIII-ompA:Ipp(脂蛋白基因) + lac(IPTG诱导转录)双启动子
pINIII-ompA:Ipp(脂蛋白基因) + lac(IPTG诱导转录)双启动子
pEZZ18:lac启动子;金黄色葡萄球菌蛋白A启动子;蛋白A的信号肽序列;两个合成的Z功能域
枯草芽孢杆菌枯草芽孢杆菌是传统的工业生产菌, 能分泌大量的胞外蛋白, 是研究蛋白质表达和分泌的良好系统,利用它进行工业规模的外源基因表达和分泌, 产生有用的蛋白类产品, 有很重要的经济价值。枯草芽孢杆菌的信号肽序列长度大多在18 到35 个氨基酸之间,并且不具有一致序列。其特定的典型结构特征是具有带正电的NH2 端, 随后是一段疏水区域(H 区) 和一个极性更强的C 区, C 区带有一致的酶切识别位点Ala-X-Ala, 酶切发生于羧基端丙氨酸之后。两个丙氨酸偶尔可以被其他具有短侧链的氨基酸代替, X 位偏爱具有巨大侧链的氨基酸。枯草芽孢杆菌信号肽中的疏水区对于蛋白质的分泌有着重要的影响, 适当的疏水性有利于蛋白的分泌, 疏水区过长或过短都不利于蛋白的分泌。有研究表明, 信号肽疏水性能提高促进青霉素G 酰化酶分泌。信号肽的选择有时能明显地影响分泌表达水平, 如枯草芽孢杆菌蛋白酶、中性蛋白酶、崖边杆菌RNA 酶( bamase) 和果聚糖酶等4 种酶的信号肽, 以芽孢杆菌BE1510 为宿主菌, 分别表达重组链霉抗生物素蛋白(streptavidin) , 结果中性蛋白酶信号肽的分泌效率最高1。
常用分泌型载体:pUB110;pE194;pUCX05-bgaB;pWB980;pHP13;pBE2;pHP13;pHP13-43(p43启动子);pHT01;pHT43;pAL12.cold-inducible secretion vector,冷诱导分泌型载体;pHT304;pMK3;pMK4;pHCMC04;pHCMC05;pMA5;pHY300PLK;pMUTIN4基因组整合型载体;pBesT502带有枯草杆菌强启动子PrepU等3。
真核分泌表达真核生物不但有细胞核、细胞质和细胞膜, 而且还有许多膜性结构的细胞器, 像原核细胞一样, 真核细胞合成的蛋白质N 端也有信号肽, 其也能形成两个α螺旋的发夹结构, 这个结构可插入到内质网膜,正在合成的多肽链带和内质网内腔内1。通常,真核表达系统比原核表达系统更能有效的表达有功能的真核蛋白,主要原因总结起来如下: 首先,原核生物缺乏真核蛋白分泌机制,使它们很难正确的表达,折叠,装配,稳定真核蛋白;其次,真核细胞与原核细胞的不同的脂质双分子层组成,以及真核细胞中更精细的转位子复合体(膜蛋白转位和插入的位点);第三,真核细胞中蛋白质翻译和折叠的速率相对较慢,这避免了新合成的蛋白质大量堆积,而大量堆积的蛋白质不利于细胞的生长代谢,有时候对宿主甚至是致命的;最后,原核生物缺乏翻译后修饰的机制,包括糖基化,磷酸化和乙酰化等,而这些机制对表达有功能活性的真核蛋白却是至关重要的。目前,主要有三个真核表达系统广泛应用于真核膜蛋白的重组表达:酵母、昆虫细胞及哺乳动物细胞,并得到了较深入的探讨和研究4。
酵母表达系统一般说来,酵母作为表达系统在真核蛋白的表达上具有一些明显的优势。和细菌一样,酵母遗传操作简单、可以在短期内大批量培养,价格低廉;同时,作为真核生物,表达的蛋白质在酵母细胞里的翻译后修饰机制与哺乳动物细胞相似。主要有两个酵母系统广泛用于蛋白质表达,它们分别是酿酒酵母表达系统和毕赤酵母表达系统。
酵母表达系统中研究较清楚、应用最广的是Pichia.Pastoris。目前酵母表达系统中常使用的信号肽包括某些外源蛋白自身的天然信号肽、α-因子信号肽( α-MF) 、蔗糖酶基因SUC2 信号肽序列、酸性磷酸酶基因PHO1 的信号肽序列、间质金属蛋白酶(MMP) /间质金属蛋白酶组织抑制剂信号肽序列。其中α-因子信号肽( α-MF) 使用最广。α-因子信号序列由87 个氨基酸组成, 是来自S.cerevsiae 的α性成熟因子前导序列, 并且已将这段序列编码的信号肽插入到几个P.Pastoris 的表达载体中( Invitrogen) , 用于外源基因的分泌表达, 如pGAPZα-A、B、C 与pMETα-A、B、C 等。在构建α-因子信号肽融合基因时, 需保留KeX2 蛋白酶切位点附近的谷氨酸- 丙氨酸(Glu-Ala) 间隔区, Glu-Ala 的存在会避免错误切割的发生1。
常用分泌型载体:pPIC9,pPIC9K,pHIL-S1,pPICZα,pYAM75P 等。
昆虫表达系统昆虫杆状病毒表达载体系统自1983 年建立以来, 已经发展成为较完善的真核表达系统, 成功的表达了多种外源基因。但信号肽在昆虫表达系统中的应用还很有限。
目前在昆虫表达载体中对信号肽在基因工程应用上的研究结果主要有: 1) 信号肽能被信号肽酶正确识别和切割, 能促进产物的分泌, 如利用α-干扰素信号肽和IL-3 信号肽构建的昆虫杆状病毒分泌载体,含蜂蜜毒肽的信号肽载体能提高木瓜蛋白酶的分泌效率5~9 倍。2) 信号肽能使产物分泌至细胞外, 但在不正确位点切割, 如利用MAG 信号肽表达的睫状体神经营养因子(CNTF)。3) 信号肽不能引导分泌表达, 如淀粉酶的信号肽不能引导木瓜蛋白酶的分泌表达。4) 信号肽不能被切除, 如用gp67 信号肽引导慈姑蛋白酶抑制剂(API) 基因在昆虫表达载体中表达虽获得了高效表达, 且表达产物的90%以上能分泌到细胞外, 但信号肽在表达过程中都没有被切除1。
哺乳动物细胞表达系统通常认为,当表达真核蛋白时,哺乳动物细胞可以为表达和维持其结构和功能完整性提供最自然的细胞环境。传统上,哺乳动物表达系统依靠的是稳定转化株的分离或者病毒载体的使用。其中使用最广泛的病毒载体是塞姆利基森林病毒(Semliki Forest Virus, SFV)。然而,细胞大批量培养及转染技术的困难和昂贵的价格限制了其在表达中的应用。近几年,随着无血清培养基及相对廉价转染试剂的发展和使用,哺乳动物细胞的大规模瞬时转染(large-scale transient transfection, LSTT)技术已经逐步被应用。欧洲结构蛋白质组学会(Structural Proteomics IN Europe, SPINE)研究了使用 LSTT法的各种蛋白质的表达特征,指出和其他表达系统相比,分泌型蛋白的表达最为完整有效,而胞质蛋白和膜蛋白则使用杆状病毒载体有更好的表达。在未来几年,在哺乳动物细胞中蛋白质表达很可能是表达体系研究中最重要的领域之一4。

 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国