

发现历程
人们早就注意到许多植物的开花具有明显的季节性,同一植物品种在同一地区种植时,尽管在不同时间播种,但开花期都差不多;同一品种在不同纬度地区种植时,开花期表现有规律的变化。
光周期现象是美国学者加奈(W.W.Garner)和阿拉德(H.A.Allard)于1920年提出的。1美国园艺学家加纳和阿拉德(GarnerandAllard)在1920年观察到烟草的一个变种(marylandmammoth)在华盛顿地区夏季生长时,株高达3~5m时仍不开花
 ,但在冬季转入温室栽培后,其株高不足1m就可开花。他们试验了温度、光质、营养等各种条件,发现日照长度是影响烟草开花的关键因素。在夏季用黑布遮盖,人为缩短日照长度,烟草就能开花;冬季在温室内用人工光照延长日照长度,则烟草保持营养状态而不开花。由此他们得出结论,短日照是这种烟草开花的关键条件。后来的大量实验也证明,许多植物的开花与昼夜的相对长度即光周期有关,即这些植物必须经过一定时间的适宜光周期后才能开花,否则就一直处于营养生长状态。光周期的发现,使人们认识到光不但为植物光合作用提供能量,而且还作为环境信号调节着植物的发育过程,尤其是对成花诱导起着重要的作用。
,但在冬季转入温室栽培后,其株高不足1m就可开花。他们试验了温度、光质、营养等各种条件,发现日照长度是影响烟草开花的关键因素。在夏季用黑布遮盖,人为缩短日照长度,烟草就能开花;冬季在温室内用人工光照延长日照长度,则烟草保持营养状态而不开花。由此他们得出结论,短日照是这种烟草开花的关键条件。后来的大量实验也证明,许多植物的开花与昼夜的相对长度即光周期有关,即这些植物必须经过一定时间的适宜光周期后才能开花,否则就一直处于营养生长状态。光周期的发现,使人们认识到光不但为植物光合作用提供能量,而且还作为环境信号调节着植物的发育过程,尤其是对成花诱导起着重要的作用。
人类早已注意到多种植物的开花时间相对稳定,但光周期在决定开花期方面所起的作用直到20世纪才了解清楚。
1912年法国J.图尔努瓦发现大麻,在每日6小时的短日照条件下会开花,在长日照下则停留于营养生长阶段。1913年德国G.A.克莱布斯发现人工加长每日照光时间,可使通常在6月开花的长春花属(Sem-pervivum)植物能在冬季开花。但明确地提出光周期理论的是美国园艺学家W.W.加纳与H.A.阿拉德。他们在1920年发现,将在美国南部正常开花的烟草(Nicotia-natabacumcv.MarylandMammoth品种)移至美国北部栽培时,夏季只长叶不开花;但如果在秋冬移入温室则可开花结实。
在北方夏季用遮光办法缩短日照时数到每天14小时以下,也可使它开花。以后发现大豆(Biloxi品种)、紫苏、高粱等也有这种现象,并各有其日长上限,日照长度短于此数值时即可开花,称此日长限度为临界日长。同时发现菠菜、萝卜等植物相反,须在日照长度超过一临界日长时才能开花。
生理效应需光种子需经长日照或连续光处理才能萌发,而嫌光种子则在短光照下与在黑暗中一样可以萌发。促进或抑制两类种子的光质要求与成花诱导的要求,都通过光敏素实现。有些植物的营养性贮藏器官的形成,也受光周期的影响。如短日照有利于马铃薯形成块茎,树木叶片的衰老脱落也与日长有关,一般在秋天短日照下容易脱落。路灯旁的行道树由于昼夜受光,落叶往往延迟。
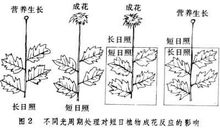
反应类型人们通过用人工延长或缩短光照的方法,广泛地探测了各种植物开花对日照长度的反应,发现植物开花对日照长度的反应有以下几种类型:2
长日植物(long-dayplant,LDP)
指在24h昼夜周期中,日照长度长于一定时数,才能成花的植物。对这些植物延长光照可促进或提早开花,相反,如延长黑暗则推迟开花或不能成花。属于长日植物的有:小麦、大麦、黑麦、油菜、菠菜、萝卜、白菜、甘蓝、芹菜、甜菜、胡萝卜、金光菊、山茶、杜鹃、桂花、天仙子等。典型的长日植物天仙子必须满足一定天数的8.5~11.5h日照才能开花,如果日照长度短于8.5h它就不能开花。
短日植物(short-dayplant,SDP)
指在24h昼夜周期中,日照长度短于一定时数才能成花的植物。对这些植物适当延长黑暗或缩短光照可促进或提早开花,相反,如延长日照则推迟开花或不能成花。属于短日植物的有:水稻、玉米、大豆、高粱、苍耳、紫苏、大麻、黄麻、草莓、烟草、菊花、秋海棠、腊梅、日本牵牛等。如菊花须满足少于10h的日照才能开花。
日中性植物(day-neutralplant,DNP)
这类植物的成花对日照长度不敏感,只要其他条件满足,在任何长度的日照下均能开花。如月季、黄瓜、茄子、番茄、辣椒、菜豆、君子兰、向日葵、蒲公英等。
除了以上三种典型的光周期反应类型以外,还有一些其他类型:
长-短日植物(long-shortdayplant)
这类植物的开花要求有先长日后短日的双重日照条件,如大叶落地生根、芦荟、夜香树等。
短-长日植物(short-longdayplant)
这类植物的开花要求有先短日后长日的双重日照条件,如风铃草、鸭茅、瓦松、白三叶草等。
中日照植物
(intermediate-daylengthplant)只有在某一定中等长度的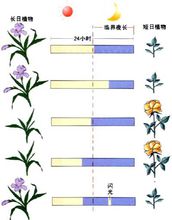 日照条件下才能开花,而在较长或较短日照下均保持营养生长状态的植物,如甘蔗的成花要求每天有11.5~12.5h日照。
日照条件下才能开花,而在较长或较短日照下均保持营养生长状态的植物,如甘蔗的成花要求每天有11.5~12.5h日照。
两极光周期植物
(amphophotoperiodismplant)与中日照植物相反,这类植物在中等日照条件下保持营养生长状态,而在较长或较短日照下才开花,如狗尾草等。
许多植物成花有明确的极限日照长度,即临界日长(criticaldaylength)。长日植物的开花,需要长于某一临界日长;而短日植物则要求短于某一临界日长,这些植物称绝对长日植物或绝对短日植物。但是,还有许多植物的开花对日照长度的反应并不十分严格,它们在不适宜的光周期条件下,经过相当长的时间,也能或多或少的开花,这些植物称为相对长日植物或相对短日植物。可以看出,长日植物的临界日长不一定都长于短日植物;而短日植物的临界日长也不一定短于长日植物。如一种短日植物大豆的临界日长为14h,若日照长度不超过此临界值就能开花。一种长日植物冬小麦的临界日长为12h,当日照长度超过此临界值时才开花。将此两种植物都放在13h的日照长度条件下,它们都开花。因此,重要的不是它们所受光照时数的绝对值,而是在于超过还是短于其临界日长。同种植物的不同品种对日照的要求可以不同,如烟草中有些品种为短日性的,有些为长日性的,还有些只为日中性的。通常早熟品种为长日或日中性植物,晚熟品种为短日植物。
生态意义在温带,温度的季节性变化幅度很大,随纬度增高而冬季延长。植物如不能在低温、霜冻到来之前形成种子,就不能繁殖后代。许多植物在长期适应过程中形成了以受气候条件影响较小的昼夜长短来感应季节的本领。例如一些起源于高纬度地区的植物,常在春季转暖之后,日照由短变长的条件下开花结实,成花要求长日照。低纬度起源的一些植物,具有短日植物特性,在日长逐渐变短的秋天开花,有利于充分利用低纬度较长的温暖时期。
在一些不是温度而是水分对植物生长起决定作用的地区,在冬季为旱季,夏季为湿季的地区起源的植物,属短日照性的较多,情况相反的地区则多为长日照植物。在同一纬度地区,对日长的不同响应使不同植物在不同季节开花结实,成为决定植物物候期的重要因素。
周期作用植物的光周期现象是指植物的花芽分化、开花、结实、分枝习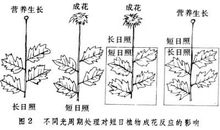 性、某些地下器官(块茎、块根、球茎、鳞茎、块茎等)的形成受光周期(即每天日照长短)的影响而言。
性、某些地下器官(块茎、块根、球茎、鳞茎、块茎等)的形成受光周期(即每天日照长短)的影响而言。
所谓光周期是指一天中,日出至日落的理论日照时数,而不是实际有阳光的时数。理论日照时数与该地的纬度有关,实际日照时数还受降雨频率及云雾多少的影响。在北半球,纬度越高,夏季日照越长,而冬季日照越短。因此,我国北方各地一年中的日照时数在季节间相差较大,在南方各地相差较小。如哈尔滨冬季每天日照只有8—9小时,夏季可达15.6小时,相差6.6—7.6小时。而广州冬季的日照时数10—11小时,夏季为13.3小时,相差2.3—3.3小时。各地生长季节特别是由营养生长向生殖生长转移之前,日照时数长短对各类药用植物的发育是个重要的因素。
植物对光周期的反应通常分为长日照植物、短日照植物、中间型日照植物三类。
长日照植物日照长度必须大于某一临界日长(一般为12—14小时以上),或者说暗期必须短于一定时数才能形成花芽,否则,植株就停留在营养生长阶段。属于这类的药用植物有红花、当归、莨菪、大葱、大蒜、芥菜、萝卜等。
短日照植物日照长度只有短于其所要求的临界日长(一般在12—14小时以下),或者说暗期超过一定时数才能开花。如果处于长日照条件下,则只能进行营养生长而不能开花。属于这类的药用植物有紫苏、菊花、苍耳、大麻、龙胆、扁豆、牵牛花等。
中间型植物这类植物的花芽分化受日照长度的影响较小,只要其它条件适宜,一年四季都能开花。属于这类的药用植物有荞麦、丝瓜、曼陀罗、颠茄等。
此外,还有所谓“限光性植物”。这种植物要在一定的光照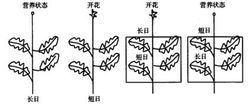 长度范围内才能开花结实。而日照长些或短些都不能开花。如野生菜豆只能在每天12一16小时的光照条件下才能开花。又如甘蔗的某些品种,它们只能在日照12小时45分钟条件下才开花。
长度范围内才能开花结实。而日照长些或短些都不能开花。如野生菜豆只能在每天12一16小时的光照条件下才能开花。又如甘蔗的某些品种,它们只能在日照12小时45分钟条件下才开花。
应当指出,植物对日照长度的反应是由营养生长向生殖生长转化的必要条件,是要在其自身发育到一定的生理年龄时才能开始,并非植物的一生都要求这样的日照长度,而绝大多数药用植物也绝不是只有一、二次的光周期处理就能引起花芽原基的分化,一般要有十几次或更多的光周期处理才能引起开花的。
临界日长是区别长日照或短日照的日照长度的标准,是指每天24小时内光照时间的多少而言,一般为12—14小时。短日照植物要求的日照时数必须短于临界日长,长日照植物要求的日照时数必须大于临界日长,否则就不能形成花芽或开花。确切的说,短日照植物并不要求较短的日照,而是要求较长的黑暗,黑暗期的长短对其花芽的形成和开花的影响更为重要。而长日照植物,光照是重要的,黑暗是不重要的,甚至是不必要的。药用植物的种类很多,对光照长短的反应差异很大,长日照与短日照之间的临界时数互相交差的情况也是可见的。
植物的光周期反应要在有一定的温度、植株的生长状态和营养条件等环境下才能进行正常发育。在影响光周期效应中,温度是个重要因素。例如,萝r、、芥菜等长日照植物,将其播种在高温长日照环境中,它们仍不能开花。这是因为高温足以抑制长日照对发育的影响。因此从事生产时,必须把光周期与温度结合起来。同理,还要与植株生长状态、营养环境等因素结合起来。
我国南北各地,由于纬度相差很大,同一生育季节里的温度、湿度 、每天的日照时数等也不尽相同,因此同一药用植物的生长发育进程也不一致,也就是说,同一药用植物在各地进入光周期的早晚或通过光周期时间长短也不相同。一般短日照植物在低纬度时进入光周期早,通过的时间也稍短,而长日照植物在高纬度下,进入光周期早,通过时间也快。另外,短日照植物,从北向南引种时,营养生长期缩短,开花结实提前。在作物、蔬菜栽培中,常利用这一特性将北方短日照品种送到南方夏种秋收,争得一茬收成。
、每天的日照时数等也不尽相同,因此同一药用植物的生长发育进程也不一致,也就是说,同一药用植物在各地进入光周期的早晚或通过光周期时间长短也不相同。一般短日照植物在低纬度时进入光周期早,通过的时间也稍短,而长日照植物在高纬度下,进入光周期早,通过时间也快。另外,短日照植物,从北向南引种时,营养生长期缩短,开花结实提前。在作物、蔬菜栽培中,常利用这一特性将北方短日照品种送到南方夏种秋收,争得一茬收成。
我们的祖先在种菜、种药及其它作物上,很早就懂得通过改变播期,调整植物生长发育时期的日照条件和温度条件,达到抑制或促进生长发育的目的。如春播萝卜(中药莱菔子)采籽,秋播萝I、收肉质根。光周期不仅影响药用植物花芽的分化与开花,同时也影响药用植物营养器官的形成。如慈姑、荸荠球茎的形成,都要求较短的日照条件,而洋葱、大蒜鳞茎的形成要求有较长的光照条件。另外,象豇豆、赤小豆的分枝、结果习性也受光周期的影响……等。
诱导机理感受光周期的部位
实验证实,感受光周期反应的部位是植物的叶片,植物成熟叶片中具有能够接受并识别光信号的光受体。迄今为止,植物中已知的光受体(色素蛋白)主要分为3 类:第1 类是光敏色素;第2 类是隐花色素;第3 类是向光素。3
实验表明,叶片感受光周期作用的能力和年龄有关:成熟叶片大于未成熟叶片;不同植物对光周期作用敏感的年龄不同:大多在子叶伸张时期;水稻在七叶期前后过了敏感期之后,年龄越大,光周期诱导的时间越长。
开花刺激物的传导
接受光周期诱导的部位是叶片,进行光周期反应的部位是茎尖的生长点,叶和起反应的部位之间隔着叶柄和一段茎。那么,必然有一个开花刺激物传导的问题。以苍耳的嫁接实验来说明。把五株苍耳植物互相嫁接在一起,且只让其中一株上的一张叶片处于苍耳开花适宜的光周期(短日照)下,其他植株都处于不适宜的光周期(长日照)下,它们都可以开花。这就证明,植株之间确有开花刺激物质通过嫁接的愈合而传递。
另外,经过短日照处理的短日植物,例如高凉菜,把其嫁接到长日植物八宝植株上,可引起八宝在短日条件下开花。反之,若将长日处理的长日植物,接到短日植物上,可引起短日植物在长日条件下开花。这说明两种光周期反应的植株所产生的开花刺激物几乎具有相同的性质。用蒸汽或麻醉剂处理叶柄或茎,可以阻止开花刺激物的运输,说明运输途径是韧皮部。苏联的柴拉轩将这种刺激物叫做成花素(forigen),但这种物质至今还没有被分离出来。
光周期诱导
光周期诱导:植物只要没得到足够日数的适合光周期,以后即便置于不适合的光周期条件下仍可开花,这种现象称作光周期诱导。4
植物完成光周期诱导的光周期处理天数(即几个光周期)因植 物而异:
物而异:
苍耳:一个光周期。即15小时照光及9小时的黑暗(15L-9D)。日本短牵牛:一天。大部分短日植物的诱导期需要一天以上,如大豆3天,大麻4天,红叶紫苏7-9天,菊12天等。长日植物需要一天的有:白芥、菠菜、油菜、毒素等。一天以上的有:天仙子:2-3天,拟南菜4天,一年生甜菜13-15天等。
不同植物光周期诱导需要的天数与植物年龄、温度、光照强度、光照长度有关。植物年龄小(达到光周期诱导的能力)、温度高、光照强,诱导期缩短。
暗期光中断现象
①光周期诱导的光照强度:在自然条件下,光周期诱导所要求的光照强度弱的,远远低于光和作用所需的光强度。一般认为在50~100勒克斯之间,有些植物甚至更低,例如水稻在夜间补充光照时,光强只需8~10勒克斯,就能明显地刺激光周期反应。说明植物光周期反应对光是极敏感的。②暗期光中断现象和生理效应暗期光中断现象表明了光周期反应与光强、光质以及参与光反应物质之间的关系。
上图结果表明:暗期对植物的开花更为重要,对于短日植物,它的开花决定于暗期的长度,只要暗期超过临界夜长(临界暗期),不管光期多长,它都开花。所以,又称短日植物为“长夜植物”更为确切。长日植物则相反,它不需要连续黑暗。假如在足以引起短日植物开花的暗期中间,被一个一定强度的闪光所间断,短日植物就不能开花,长日植物就能开花,这种现象叫做暗期光中断现象。
用不同波长的光来间断暗期的试验表明:无论是抑制短日植物开花,还是诱导长日植物开花,都是红光最有效。如果在红光照过后立即再用远红光照,那么暗期闪光间断的效应就会消失。说明暗期闪光间断效应有光敏素参与。
光敏素及其在成花中的作用
暗期闪光中断效应作用的最大作用光谱正好是光敏素的最大吸收光谱,最小作用光谱也正好是光敏素的最小吸收光谱。①光敏素的物理化学性质
光敏素可以从高等植物几乎所有部分中提取出来,也就是说存在于高等植物的所有组织(根、茎、叶、花、果、实、种子、胚芽鞘)中。在细胞中,光敏素可能集中在细胞的膜的表面上。绿色组织中光敏素含量较低,在黄化组织中较高,浓度级为10-7—10-5M。
高纯度的光敏素已从黄化的单子叶植物幼苗(玉米等)中制的。经鉴定是一种蓝色蛋白质,它的生色团类似叶绿素和血红素,有四个吡咯环,但它们不是环状连接,而是开放 成直链,以共价键与蛋白质部分相连,即由蛋白质和生色团两部分所组成。天然的光敏素分子量大约为120kDa。
成直链,以共价键与蛋白质部分相连,即由蛋白质和生色团两部分所组成。天然的光敏素分子量大约为120kDa。
②光敏素与开花诱导
光敏素有两种存在形式,Pr和Pfr。在黄化组织中,光敏素大部分以红光吸收型(Pr)存在,其吸收高峰在660nm。当用红光照射时,Pr的吸收光谱发生变化,吸收高峰在725nm。这说明通过红光照射Pr已转变为另一种形式(Pfr)。
光敏素在开花中如何产生生理效应,当前还不清楚。光敏素本身并不是开花刺激物,但它可以触发开花刺激物的形成(合成或激活)。当前一般认为,不论是短日植物(SDP)还是长日植物(LDP),其开花都与Pfr与Pr的比例有关。对于SDP,在光期结束时,Pfr/Pr比值高(因为在白天,红光比例大,有利于Pfr的形成),开花刺激物的合成受到阻止。转入夜间后,Pfr向Pr逆转,Pfr/Pr比值变小。当此值到一定水平时,就会触发引导开花刺激物形成的代谢过程,SDP的成花反应就可以发生。如暗期为红光所间断,Pr转换成Pfr,Pfr/Pr比值升高,开花刺激物的形成即遭受阻止。
对于LDP,形成开花刺激物,要求较高的Pfr/Pr比值,这一比值可在长光期结束时获得。如果在短日照下,暗期过长,则Pfr转变为Pr,或Pfr受到破坏,Pfr/Pr值达不到较高水平,开花刺激物就不能形成。暗期的红光间断,可使Pfr/Pr值重新提高,使开花刺激物得以合成,长日植物就可以开花。
光周期刺激的感受和传递
植物在适宜的光周期诱导后,发生开花反应的部位是茎顶端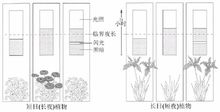 生长点,然而感受光周期的部位却是植物的叶片。若将短日植物菊花全株置于长日照条件下,则不开花而保持营养生长;置于短日照条件下,可开花;叶片处于短日照条件下而茎顶端给予长日照,可开花;叶片处于长日照条件下而茎顶端给予短日照,却不能开花(图9-10)。这个实验充分说明:植物感受光周期的部位是叶片。对于光周期敏感的植物,只有叶片处于适宜的光周期条件下,才能诱导开花,而与顶端的芽所处的光周期条件无关。虽然也有少数植物的其它部位对光周期有一定的敏感性,如组织培养的菊苣根可对光周期起反应,但感受光周期最有效的部位是叶片。叶片对光周期的敏感性与叶片的发育程度有关。幼小的和衰老的叶片敏感性差,叶片长至最大时敏感性最高,这时甚至叶片的很小一部分处在适宜的光周期下就可诱导开花。例如,苍耳或毒麦的叶片完全展开达最大面积时,仅对2cm2的叶片进行短日照处理,即可导致花的发端。
生长点,然而感受光周期的部位却是植物的叶片。若将短日植物菊花全株置于长日照条件下,则不开花而保持营养生长;置于短日照条件下,可开花;叶片处于短日照条件下而茎顶端给予长日照,可开花;叶片处于长日照条件下而茎顶端给予短日照,却不能开花(图9-10)。这个实验充分说明:植物感受光周期的部位是叶片。对于光周期敏感的植物,只有叶片处于适宜的光周期条件下,才能诱导开花,而与顶端的芽所处的光周期条件无关。虽然也有少数植物的其它部位对光周期有一定的敏感性,如组织培养的菊苣根可对光周期起反应,但感受光周期最有效的部位是叶片。叶片对光周期的敏感性与叶片的发育程度有关。幼小的和衰老的叶片敏感性差,叶片长至最大时敏感性最高,这时甚至叶片的很小一部分处在适宜的光周期下就可诱导开花。例如,苍耳或毒麦的叶片完全展开达最大面积时,仅对2cm2的叶片进行短日照处理,即可导致花的发端。
昆虫学
对昆虫滞育产生影响的光周期(Photoperiod)是指一昼夜中的光照时数与黑暗时数的节律,一般以光照时数表示,是影响昆虫滞育的主要因素。引起昆虫种群中50%的个体滞育的光照时数,称为临界光周期(critical photoperiod)。不同种或同种不同地理种群的昆虫,其临界光周期不同,如三化螟南京种群为13小时30分,广州种群为12小时。感受光照刺激的虫态,称为临界光照虫态。临界光照虫态常常是滞育虫态的前一虫态,如家蚕以卵滞育,其临界光照虫态为上一代成虫;亚洲玉米螟以老熟幼虫滞育,其临界光照虫态为3~4龄幼虫;棉铃虫以蛹滞育,其临界光照虫态为1~5龄幼虫。处于临界光照虫态的昆虫对光的反应极为敏感,一般只需l~2Lx(勒克斯)的照度就能发生作用,如钻入苹果内的梨小食心虫,当果面的光照度只有1~3Lx时,即能对光作出反应。根据昆虫滞育对光周期的反应,可将昆虫分为以下4种滞育类型。
1、短日照滞育型 即长日照发育型。其特点是昆虫滞育的个体数随日照时数的减少而增多。通常光周期长于12~16小时,仍可继续发育而不滞育。一般冬季滞育的昆虫,如亚洲玉米螟等属于此类型。
2、长日照滞育型 即短日照发育型。其特点是昆虫滞育的个体数随日照时数的增加而增多。通常光周期短于12小时,仍可继续发育而不滞育。一些夏季滞育的昆虫,如小麦吸浆虫等属于此类型。
3、中间型光周期过短或过长均可引起滞育,只有在相当窄的光周期范围内才不滞 育。如桃小食心虫在25℃时,光照短于13小时,老熟幼虫全部滞育;光照长于17小时,半数以上滞育;而光照为15小时时,则大部分不滞育。
4、无光周期反应型 光周期变化对滞育没有影响。如苹果舞毒蛾、丁香天蛾等。
控制应用光周期应用
对于那些对光周期敏感的植物,认识其对光周期响应的类型,对于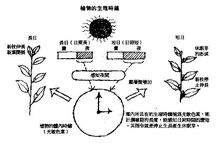 掌握和控制其开花、结实等生长发育时期在一年中的分配,具有重要意义。例如:
掌握和控制其开花、结实等生长发育时期在一年中的分配,具有重要意义。例如:
①引种时要详细了解当地日长的季节变化和待引品种的光周期要求。如果日长条件不符合其要求,则不能开花或开花延迟,种子不能成熟;过早地满足成花要求,又会不必要地缩短生长季,对干物质的累积不利。
②人工缩短或延长光照时间可以促进或延迟开花,甚至可以使许多原来开花季节较固定的花卉在任何季节开花。在杂交育种中可以用人工长日或短日处理来使父母本花期相遇,提高杂交种子产量。
③在育种工作中,当作物种子在收获后立即播种,而光周期不利于开花结实时,人工调整日长,可使之及时成花、结实,从而加速世代繁育,缩短育种年限。中国育种学家创造了南繁北育的办法,利用异地温度和光周期的条件,使某些作物可一年繁育2~3代。例如:中国水稻育种家常利用海南岛冬季日照短、较温暖来进行冬繁,使良种繁育时期缩短。
④对收获营养体的作物,如烟草、洋麻、黄麻、亚麻等,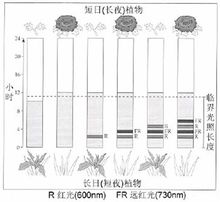 移至北方栽培,利用长日条件推迟或防止成花,可提高烟叶或麻纤维的产量或品质。
移至北方栽培,利用长日条件推迟或防止成花,可提高烟叶或麻纤维的产量或品质。
利用对植物光周期特性的知识控制生长发育的办法,大体上可分为两类:
①人工改变日长。因为成本较高,多用于经济价值较高的花卉的栽培或作物品种的繁育。其中光间断打断长夜所需光量较小,比人工照光延长光照时间经济得多。
②选择品种特性,以适应特定地区自然光照特点。这可用于多种作物。为了使所得品种能推广于较大的纬度范围,近代育种工作者有选育对光周期较不敏感的遗传特性的趋势。
光周期在农业生产上的应用
1、引种:纬度相近地区引种易成功。短日照植物北移因生长季日照延长,长日照植物南移因生长季日照缩短,都有延迟发育的作用;反之,短日照植物南移,或长日照植物北移有促进发育的作用。短日照植物引种时,温度和光照长度的效应是相互叠加的,对发育期提早或推迟 的影响较为突出,南北距离较远时,则不易成功。长日照植物南北引种,光温影响是补偿的,一般较易成功。但在热量条件较差地区,从高纬度引种短日照植物,往往有利于避霜早熟。但对收获营养体为主的作物,则要防止过早向生殖生长转化。
的影响较为突出,南北距离较远时,则不易成功。长日照植物南北引种,光温影响是补偿的,一般较易成功。但在热量条件较差地区,从高纬度引种短日照植物,往往有利于避霜早熟。但对收获营养体为主的作物,则要防止过早向生殖生长转化。
2、育种工作中利用光周期调节花期实现品种间杂交。利用光周期效应使作物提早发育,还可以实现加代繁殖缩短育种年限。
3、调节播栽期。选择适宜的播种期和移栽期,使自然日照、温度与作物的要求相一致,是提高产量的主要条件。
4、花卉栽培。利用光周期现象进行人工调节花期,能使短日照植物的菊类在任何季节开花。
光强要求
诱导成花对光周期中光暗时间的长短和间隔的要求,有时很严格,但所需光强却很低,而且光强进一步提高并不影响成花响应。但如果要以短时间照光引起光间断,则需较强的光。
在可见光中,光照诱导成花或光间断破坏诱导的效应,以660纳米的红光为最有效。这种作用可以被730纳米的远红光解除,又可以被660纳米的红光恢复。这种相互逆转可以反复多次。这表明接受光周期信号的色素是植物。660纳米的红光将光敏素转变为生理上活跃的远红光型,而730纳米的远红光则把远红光型转变为生理上不活跃的红光型(Pr)。在黑暗中的衰变速率和向Pr的转化速率,可能对光周期响应的计时起一些作用。但的暗衰败速率随温度升降变化很明显,而有些植物的光周期长短却突出地不受温度变化的影响,所以植物体内必定还另有确定节奏的生物钟。它的机理,至今仍不清楚。有一些植物的光周期响应在高温下与低温下不同。例如短日植物中的牵牛、高凉菜、苍耳等,在低温条件下,即使在长日照下也可成花。长日植物中的二色金光菊(Rudbeckiabicolor)、风轮菜属植物(Calaminthaofficinalis)在高温处理下,Blitumcapitatum与B.viratum在低温下均可在短日照下开花。
刺激感受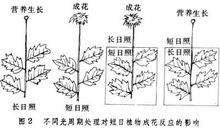
光周期诱导成花过程在解剖上最早看到的是苗端开始分化花芽,但是否分化花芽取决于叶片是否处于适当的光周期条件下。可见感受光周期的部位不是苗端,而是叶片。有些对光周期敏感的植物,只要一片叶或甚至一片叶的一部分(例如苍耳只要长成叶的2平方厘米)接受合适的光周期,即使其余部分都处在不利于成花的光周期下,也能分化花芽。显然,接受光周期的叶片,必须以某种方式把光周期诱导的信号传递给苗端的分生组织。
叶片所接受的光周期刺激,不但可以传递给本枝条,而且在本枝条去顶芽后,还可以传递给同株的另一个去掉了叶片的枝条的顶芽,使它分化出花芽。当把经过诱导的植株的叶片嫁接到另一株未经诱导的植株上去以后,也可以诱导它成花。从后一植株取下的叶片,又可以嫁接的方式诱导另一株未经诱导的植株成花。这种嫁接在苍耳上可以辗转5~6次,仍然生效。
根据这类实验结果,苏联M.X.柴拉希扬于1937年提出,诱导叶中形成一种物质,传导到顶芽分生组织后引起成花,他称这种物质为成花素。但经过几十年的努力,还未能分离和鉴定出这种物质。
另一些植物,如草莓、藜(短日植物)和天仙子(长日植物),只要将全部叶子去掉(新叶随生随除),在任何日长下都能开花。表明在这些植物的叶子里,不是在适宜的日照下形成促进花芽分化的物质,而是在不适宜的日照下形成某种抑制花芽分化的物质,称为成花抑制物,所以去掉叶片反而容易开花。
对花影响无论长日植物或短日植物,一般只要求经历一定天数的长日或短日,就能成花,并不要求长期处于那种日照长度之下。有些短日植物,如苍耳,甚至一个短日长夜周期,即可成花。但从这一合宜的光周期处理到花芽分化,还需一段时期。这种适合的光暗周期使植物从不分化花芽的营养生长阶段转入分化花芽的状态的作用,称为光周期诱导。
长日植物所需的日照长度因植物种类而异。如一年生甜菜为13~14小时,冬小麦为12小时,天仙子为10小时,并不一定要比12小时长。短日植物所需的临界日长多数短于12小时,但也有长于12小时的,如某些大豆品种为13.5小时,苍耳为15.5小时。所以在昼长13小时夜长11小时的光周期下,长日植物冬小麦、天仙子和短日植物晚大豆(Biloxi)、苍耳都可以开花。可见,长日植物与短日植物的区别,不在于临界日长是否大于12小时,而在于要求日长大于还是小于某临界日长。

无论对于短日植物或长日植物,以光将长夜打断,都产生类似于短夜的效应(短日植物不能成花,长日植物能成花);而以暗间断将长日打断,却不影响长日的效应(短日植物仍不能成花,长日植物仍能成花)。所以关键是夜长而非昼长。但夜长也不是决定能否诱导成花的唯一因素。如只需一天短日长夜的藜,在连续光照条件下,给以72小时黑暗处理,并在这个暗期内的不同时间做光间断处理,如果在暗期开始3小时后照射2分钟红光,则虽然以后的暗期还有69小时,远远大于临界暗期,植物也不能成花。而如果在暗期开始后18小时做光间断处理,则植株不但能开花,而且成花数比一个通常的短日长夜处理还多。光间断效应的大小与正负,与它在光周期中的时刻有关。按E.比宁的理论,许多植物(以及动物)体内有一种内在节奏,称为生物钟或生理钟。它的周期接近但不等于24小时,一般在21与28小时之间。这个周期称为近似昼夜周期。在周期的不同相位上,植物对光或暗处理的敏感性不同。
影响因素光周期反应是影响作物由营养生长向生殖生长生理转变过程的重要因素,是一种受环境影响的复杂网络调控,多基因控制及多基因间相互协调和制约决定了调控的复杂性。研究发现,多个参与光周期反应的基因且这些基因在一定程度上影响光周期敏感性。我国地域辽阔,南北跨度大,具有热带、亚热带和温带等多种热量带,环境条件复杂多变。作物通过形态和生理生化变化协调自身生长发育以适应不同的环境条件。遗传和环境互作使相同种质在不同环境条件下表现不同。
光周期特性与地域生态条件存在一定关系。在不同的种植区域,适宜的小麦品种利用充分的光照资源,能够最大限度地发挥品种的潜力,从而提高产量。在低纬度地区,光周期不敏感型品种有利于小麦赶在夏季高温来临之前完成灌浆;而在高海拔地区,光周期敏感型品种可推迟开花,防止小麦过早形成花原基而避免早春霜冻的伤害。国内有研究认为,不同麦区在育种和栽培实践中应有不同的育种策略:西南麦区可以考虑使用光周期敏感型品种,充分发挥低温短日照优势;华南冬麦区需要考虑使用对温度敏感的品种,避免使用光周期敏感型品种;东北春麦区则需要推广使用光周期敏感型品种,以提高该区小麦品种的光周期敏感性。5


 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国