

形态特征
马兜铃猪笼草为藤本植物。茎可分枝,可长达8米,直径约5毫米,呈圆柱形至钝棱柱形。节间距可长达15厘米。叶芽明显突出,位于叶腋上1.5至7毫米处。
在马兜铃猪笼草的自然分布地中已发现了小的变种,但尚未有关于其种下类群的描述。
叶片革质,无柄。呈汤匙形至披针形,可长达15cm,宽至3cm。叶片末端尖细,基部缓缓缩小。各有2~5条纵脉从中脉分出。羽状脉不明显。笼蔓长达15cm。
马兜铃猪笼草的下位笼只会产生于还未开始拉长攀爬的茎和攀援茎的侧芽。下位笼下三分之二为漏斗状,上三分之一为球形。可长达7cm,宽至3cm。在每个笼子的前部有一对小于4mm的翼。唇可达5mm,向内弯曲。在唇的内缘,有一排排弯曲的小唇齿。捕虫笼的内表面几乎全部覆盖着腺体。腺体凸出,直径为0.2~0.3mm,密度约为每平方厘米200~500个。捕虫笼的盖子为圆形或心形,约1.5cm宽,没有附属物。在盖子基部的后方有一个分叉或不分叉的笼蔓尾,长度小于7mm。
上位笼为漏斗形,上半部类似囊状。上位笼比下位笼大,可高达15cm,宽至8cm,捕虫笼前部的翼缩小为一对凸起。笼口几乎与地面垂直。唇可达6mm,向内弯曲。在唇的内缘,有一排排弯曲的小唇齿。捕虫笼的盖子为卵形。捕虫笼内表面下部的三分之二覆盖着腺体。腺体略微凸出,直径0.2~0.4mm,密度为每平方厘米200~250个。在其他很多方面,下位笼与下位笼很相似。
马兜铃猪笼草的花序为总状花序。花梗和花轴都可以达到15cm长,通常雌株较短些。小花梗带小苞片与一朵花,可长达12mm。萼片呈卵形,可长达4mm。果荚可长达20mm,宽至4mm,呈披针形。种植为丝状。
马兜铃猪笼草植株大部分无毛被。在叶腋、中脉和捕虫笼有稀疏的白色短毛。
茎和叶片都为淡绿色。捕虫笼为白色或淡红色带红褐色斑点。唇为暗红色。盖子的整个下表面为红色,上表面带斑点。干燥标本为棕色,捕虫笼上保留有黑色斑点。
马兜铃猪笼草还未发现变种或个体。
|| ||
生长环境生长于海拔1800米至2500米的地区。
分布范围苏门答腊
植物学史马兜铃猪笼草首次由威廉·梅哲(Willem Meijer)于1956年8月5日在占碑的土朱山(Mount Tujuh)发现的。它的编号为Meijer 6542的模式标本收集于土朱山海拔2000m的地区。现存放于荷兰莱顿的荷兰国家植物标本馆(the National Herbarium of the Netherlands)。除此之外,另一个副本存放于爪哇的茂物植物园(the Bogor Botanical Gardens )中。
虽然它被标记为“新物种?”,但它还是被忽略了几乎30年。直到1988年,约阿希姆·那兹(Joachim Nerz)访问莱顿大学植物标本馆(the herbarium of Leiden University)后才被注意到。1994年,这个物种才被命名为马兜铃猪笼草(N. aristolochioides)。同年,分类学家简·斯洛尔(Jan Schlauer)研究了编号为Meijer 7426的模式标本后,错误的认为马兜铃猪笼草看起来垂直的笼口是因为标本压制不当造成的。
1996年,那兹在法兰克福棕榈公园(Frankfurt Palmengarten)遇见了梅哲,在那梅哲给那兹看了一种神秘物种的照片。之后,1996年6月,连同卡特琳·欣德·霍弗(Katrin Hinder hofer),那兹组织了一次苏门答腊实地考察,再次在野外找到了马兜铃猪笼草。
1997年,马修·杰布(Matthew Jebb)和马丁·奇克(Martin Cheek)正式描述了马兜铃猪笼草,并发表在一本植物学杂志上。第二年,约阿希姆·那兹(Joachim Nerz)也写了一篇关于马兜铃猪笼草的详细描述,发表在《食虫植物通讯(Carnivorous Plant Newsletter)》上。
|| ||
生态学
 马兜铃猪笼草生长于陡峭的山脊上的森林中。它通常扎根于地面,也可以附生于树干上的水苔中。它的海拔分布范围是1800~2500m。
马兜铃猪笼草生长于陡峭的山脊上的森林中。它通常扎根于地面,也可以附生于树干上的水苔中。它的海拔分布范围是1800~2500m。
2001年,查尔斯·克拉克(Charles Clarke)认为马兜铃猪笼草只存在于占碑(Jambi)的土朱山(Mount Tujuh)。而之后赫伯特·斯托弗·罗宾逊(Herbert Christopher Robinson)和塞西尔·博登·克洛斯(Cecil Boden Kloss)收集的标本来自葛林芝山(Mount Kerinci),所以他们认为马兜铃猪笼草可能分别得更为的广泛。葛林芝山位于土朱山附近,是苏门答腊的最高峰。克拉克认为葛林芝山大部分尚未开发,马兜铃猪笼草很有可能会在这有分布。关于马兜铃猪笼草在土朱山的分布了解得也不是很全面,因为在土朱山的七个山峰中,只考察了其中的三个。
2009年,斯图尔特·麦克弗森(Stewart McPherson)了解到,马兜铃猪笼草在葛林芝·塞布拉国家公园(Kerinci Seblat National Park)原有三处原生地,但其中的一处可能已经消失了。马兜铃猪笼草原生地周围同时存在着裸瓶猪笼草(N. gymnamphora )和欣佳浪山猪笼草(N. singalana )。马兜铃猪笼草生长于山地森林和火山湖沿岸长满露兜树(Pandanus)的沼泽中。裸瓶猪笼草(N. gymnamphora )的海拔分布范围是1800~2100m,虽然与马兜铃猪笼草的原生地有部分重叠,但没有发现它们之间的自然杂交种。个头较小的欣佳浪山猪笼草(N. singalana )与马兜铃猪笼草的分布范围一致。但它们似乎占据了不同的生态位置,欣佳浪山猪笼草(N. singalana )通常在地表生活,而马兜铃猪笼草则常攀爬至树冠上。马兜铃猪笼草与欣佳浪山猪笼草(N. singalana )的自然杂交种已被发现。
马兜铃猪笼草已被列入《2006年世界自然保护联盟红色濒危物种名录(2006 IUCN Red List of Threatened Species)》中,等级为极危(Threatened)。在做评估的时候它只在一座山上有分布了。尽管它所有的分布都位于葛林芝·塞布拉国家公园(Kerinci Seblat National Park)中,但它依然因为其独特的捕虫笼外表而受到过度采集的威胁。
食虫性 马兜铃的下位笼常生长于茂密的水苔上,它的笼口位于捕虫笼的顶部。约阿希姆·那兹(Joachim Nerz)认为,下位笼只是捕食地面昆虫的。而上位笼却很特别,和克罗斯猪笼草(N. klossii )一样,马兜铃猪笼草的上位笼上也有白班,使得阳光可以照入捕虫笼内。这样从捕虫笼的内部看,唇和盖子的部分就会显得很黑暗,而笼身上的白斑就会显得很明亮,飞虫在捕虫笼中就会误以为白斑处是出口而困在捕虫笼内,最终筋疲力尽落入消化液中。这个结构类似北美的一些瓶子草的结构,如眼镜蛇瓶子草(Darlingtonia californica),小瓶子草(Sarracenia minor)和鹦鹉瓶子草(Sarracenia psittacina)。在贾桂林猪笼草(N. jacquelineae)身上也有类似的机制。
马兜铃的下位笼常生长于茂密的水苔上,它的笼口位于捕虫笼的顶部。约阿希姆·那兹(Joachim Nerz)认为,下位笼只是捕食地面昆虫的。而上位笼却很特别,和克罗斯猪笼草(N. klossii )一样,马兜铃猪笼草的上位笼上也有白班,使得阳光可以照入捕虫笼内。这样从捕虫笼的内部看,唇和盖子的部分就会显得很黑暗,而笼身上的白斑就会显得很明亮,飞虫在捕虫笼中就会误以为白斑处是出口而困在捕虫笼内,最终筋疲力尽落入消化液中。这个结构类似北美的一些瓶子草的结构,如眼镜蛇瓶子草(Darlingtonia californica),小瓶子草(Sarracenia minor)和鹦鹉瓶子草(Sarracenia psittacina)。在贾桂林猪笼草(N. jacquelineae)身上也有类似的机制。
在马兜铃猪笼草的捕虫笼内并没有发现存在共生生物。但这不是由于这个区域不存在这类共生生物,因为在同区域生长的欣佳浪山猪笼草(N. singalana )的捕虫笼内就有大量的共生生物。有学者认为,这可能是因为马兜铃猪笼草的捕虫笼结构也会使那些共生生物迷失方向,所以那些共生生物不会选择在马兜铃猪笼草的捕虫笼中生存。
马兜铃猪笼草的上位笼和下位笼具有不同的捕食机制。
陷阱机制马兜铃的下位笼常嵌入茂密的水苔中,仅其顶部可见。约阿希姆·那兹认为下位笼仅捕食地面昆虫。昆虫爬到捕虫笼顶部的笼口时就可失足落入捕虫笼内而无法爬出,最后被消化液淹死。
光诱机制克洛斯猪笼草(N. klossii)和马兜铃猪笼草是猪笼草属中仅有的两种具有带白斑球状部分捕虫笼的物种,阳光可透过白斑照入捕虫笼内。从捕虫笼的内部看,唇和盖子较黑暗,而笼身球状部分的白斑则较明亮。类似的结构还存在于北美的一些瓶子草上,如眼镜蛇瓶子草(Darlingtonia californica),小瓶子草(Sarracenia minor)和鹦鹉瓶子草(Sarracenia psittacina)。贾桂琳猪笼草(N. jacquelineae)也具有类似的捕虫机制。 事实证明,马兜铃猪笼草所捕捉到的猎物都是由飞虫组成的,其都具有趋光性。飞虫在捕虫笼中就会误以为白斑处是出口而困在捕虫笼内,最终落入消化液中。因此,马兜铃猪笼草的捕虫笼有类似龙虾笼的功能。1
马兜铃猪笼草以半透明穹顶为核心的捕虫机制已得到了实验数据的支持。在2012年一项对马兜铃猪笼草光诱机制的研究中,用红色赛璐珞覆盖捕虫笼顶部的捕虫笼其果蝇捕获率比正常捕虫笼减少了约三分之二(果蝇为红色盲,仅对紫外线、可见光内的蓝绿波段敏感)。
粘捕机制马兜铃猪笼草可产生大量粘稠的消化液,并在捕虫笼的内表面上形成一层薄膜。至少马兜铃猪笼草的上位笼已发展出了利用粘稠的内表面来捕捉飞虫的功能。其他七种与具密切近缘关系的苏门答腊猪笼草物种也同样具黏稠消化液,分别为疑惑猪笼草(N. dubia)、杏黄猪笼草(N. flava)、无刺猪笼草(N. inermis)、贾桂琳猪笼草、马桶猪笼草(N. jamban)、塔蓝山猪笼草(N. talangensis)和细猪笼草(N. tenuis)。与马兜铃猪笼草一样,这些物种都具漏斗形的捕虫笼,且其内表面几乎都完全覆盖着消化腺。2
相关物种马兜铃猪笼草的捕虫笼与其他猪笼草有着巨大的差别。它的笼口几乎与地面垂直。
约阿希姆·那兹(Joachim Nerz)认为马兜铃猪笼草与塔蓝山猪笼草(N. talangensis)有着近缘关系。但通过笼口的特点可以容易的将它们区分开来。塔蓝山猪笼草(N. talangensis)的笼口是与地面水平的。此外,塔蓝山猪笼草(N. talangensis)有一个很短的唇颈,而马兜铃猪笼草缺少这个结构。无论是笼口还是盖子,塔蓝山猪笼草(N. talangensis)的都较大。它们的生长习性也不同,塔蓝山猪笼草(N. talangensis)只生活于地面,不善于攀爬,而马兜铃猪笼草则会常常作为一个附生植物,攀爬到林冠上。
2001年,查尔斯·克拉克对来自苏门答腊岛和西马来西亚的猪笼草进行了分支系统学分析,共利用了70个形态特征。以下为猪笼草进化树的“第1分支”,其相关性达51%。3
以下是得到的进化树的一部分。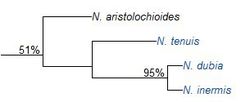
虽然马兜铃猪笼草在很多方面与克罗斯猪笼草(N. klossii )很相似,但这两个物种间存在地理隔离,所以不认为他们的关系很近。这只是一种趋同进化的表现,即是不紧密相关的两个独立的物种却获得相似特征的现象。
自然杂交种目前发现的马兜铃猪笼草的自然杂交种只有一种,即是马兜铃猪笼草与欣佳浪山猪笼草(N. singalana )的自然杂交种。它存在于土朱山的两个山脊上的水苔森林中,在其中一个山脊上有马兜铃猪笼草分布。这种自然杂交种是很罕见的,因为马兜铃猪笼草与欣佳浪山猪笼草(N. singalana )的花期并不相同。它体型比父母本都要小,捕虫笼很少会超过5cm高。它的下位笼与塔蓝山猪笼草(N. talangensis)很相似,但唇齿更发达。上位笼的下部为漏斗形,中部为球状,上部为圆柱形。这个自然杂交种区别于马兜铃猪笼草的特点是它的唇更窄且为圆柱形,笼口倾斜,而不像马兜铃几乎与地面垂直。3

 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国