

鸭嘴龙类Hadrosauridae 为栖息于白垩纪后期的草食恐龙,属于爬行纲鸟龙目鸟脚亚目的一种。萨哈林岛(库页岛)产的日本龙(Nipponosaurus sachalinensis)也属于这一种。头骨前后长,上颌和下颌都宽扁。日本名就是因口形而命名的。多数都具有紧密排列的菱形齿,总数达二千个,象磨那样最适于粉碎食物。
简介鸭嘴龙类是鸟脚类恐龙的一个属群头骨较长,上颌和下颌有些宽扁,多数都具紧密排列的菱形齿,就像磨一样,适于粉碎物。它们主要用两脚行走,有时可能会用四行走。鸭嘴龙类恐龙经常涉入近海岸的浅域,以便寻觅水生植物,有时则在陆上寻找物,万一遇到大型肉食性恐龙就赶快逃入中。因而鸭嘴龙类的化石也一般发现于河流湖泊、海岸和浅海等处的沉积层中1。
蒙古高原特有的鸭嘴龙类巴克龙是蒙古高原特有的鸭嘴龙类。它头骨短而平滑,牙齿较少并呈棱柱形。其臼齿达,能有效地将植物磨碎消化。巴克龙还有特点,就是旧的牙齿磨蚀后,不断会有新的牙长出来补充。巴克龙的前肢较短,后肢长而壮,脊骨处有不寻常的大尖刺突出。它们平常两脚行走,生活于河湖附近,以植物为食。成的巴克龙有6米长,四脚站立时可达到2米高,重在1.3吨左右1。
鸭嘴龙类恐龙鸭嘴龙类恐龙是白垩纪晚期最繁盛的恐龙。科学家根据已发现的化石,把它分成2个亚科,36个属。像我们所熟知的壮龙、粗齿龙等,都属于鸭嘴龙类。鸭嘴龙类恐龙吻端宽而扁平,状如鸭嘴,因而得名。它们一般身体壮大,长约10余米,重数吨。它们通常两足行走,但有时四足行走。它们前肢短于后肢,两者之比约为1:1.6。前肢拇指缺失,第二至第四指发育,第三指最发育,指端有蹄状爪,第五指细小。后肢第第五趾退化,仅有3趾。它们头长,前上颌骨及前齿骨无牙齿,颊部菱形细齿密集排列,有3~5排牙齿依次替换,同时可有1~3排牙齿承担咀嚼功能。它们有的是平头,有的头上生有顶饰(又称冠状嵴)。
鸭嘴龙类分为壮龙亚科和兰博龙亚科。壮龙亚科有15个属,它们多是平头,如有顶饰,顶饰也是实心的。它们的荐椎具腹沟,荐神经棘长宽比小于4.5中国的山东龙就属于这个亚科。兰博龙亚科有8个属,它们头上具中空的顶饰,荐神经棘长宽比大于4.5,其前肢比壮龙稍短而且更粗壮2。
物种解释推断体长约10米,体重约4吨。主要是两脚行走,可能有时也用四脚行走。它们可能有时涉入近海岸的浅水域,挖掘水底寻食水生植物,有时在陆上寻找食物,在遇到大形肉食恐龙时便逃入水中。化石在河流、湖泊、海岸和浅海等处的沉积层中发现。有时具有皮革样的皮肤和木乃伊的感觉,表现有指间有蹼和脊柱上的长腱使象橹那样的活动。头骨鼻的附近也具有多种适应形态,前颌骨和鼻骨呈盔状棱角的Corythosaurus,后头部有长梭角的Parasaurolo就是明显的例子。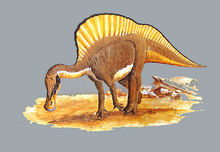
早期研究水栖鸭嘴龙类
最早被发现、鉴定的鸭嘴龙类化石,是在1956年发现的糙齿龙牙齿化石,以及在1958年发现的鸭嘴龙属化石,包含脊椎、骨盆、后肢、数颗牙齿、破碎的颌部骨头。这些鸭嘴龙类化石都没有完整的头颅骨。雕刻家本杰明·瓦特豪斯·郝金斯(Benjamin Waterhouse Hawkins)在重建鸭嘴龙属的头骨过程中,曾参考鬣蜥的头部。约瑟夫·莱迪(Joseph Leidy)借由其他部分的化石,以做出鸭嘴龙属的古生物学研究。莱迪认为它们的前肢、后肢长度相近,因此推测鸭嘴龙属是种外表类似科莫多龙的四足爬行动物,是生存在河岸、水陆两栖的动物,以前肢勾取植物为食。爱德华·德林克·科普(Edward Drinker Cope)支持莱迪的假说,并提出鸭嘴龙类的牙齿与颌部关节结构虚弱,因此只能以质地柔软的水生植物为食。
在1874年,科普在科罗拉多州发现了柱齿龙的化石,化石仍然破碎、不完整,但与鸭嘴龙属相比,柱齿龙的颌部骨头比较完整,并首次呈现出鸭嘴龙类的复杂齿系(Tooth batteries)。在1882年,J. L. Wortman与R. S. Hill发现了一个部分骨骼,其中包含完整的头颅骨,他们将这个化石交给科普。在1883年,科普将这标本建立为双芽龙的一个新种,奇异双芽龙(D. mirabilis);经过多次的重新归类后,这个标本已成为大鸭龙的正模标本。科普注意到头颅骨的前段长而宽、形状低矮。科普形容,大鸭龙的头部侧面类似天鹅,从上方看则类似琵鹭。科普也发现这头颅骨的喙状嘴,似乎有皮肤组织的痕迹。经过研究之后,科普仍将鸭嘴龙类叙述成生活在岸边、水陆两栖的动物,只能以水生植物为食。科普的理由是鸭嘴龙类的颌部关节、下颌牙齿虚弱,它们若以质地坚硬的陆地植物为食,颌部关节、下颌牙齿可能会脱落。但是,科普当时取得的标本并不完全,缺少颌部内侧、支撑牙齿的骨头,鸭嘴龙类的牙齿其实非常牢固。科普曾尝试叙述整个头骨与骨骼,但研究并没有完成,就已死去。
在科普死后,他的长期竞争对手奥塞内尔·查利斯·马什(Othniel Charles Marsh)进行破碎龙的研究工作。马什修正科普对鸭嘴龙类的数项特征描述,但仍抱持鸭嘴龙类以柔软的水生植物为食的理论。直到1922年,在一个鸭嘴龙类化石发现疑似胃内含物组成,由陆地植物所组成,例如:针叶树的树枝、树叶、水果、种子,该研究人员做出新假设,鸭嘴龙类同时以陆地植物、水生植物为食。
1940年代:理论转变
在1942年,理查·史旺·鲁尔( Richard Swann Lull)与尼尔达·莱特(Nelda Wright)提出一份鸭嘴龙类专题论文,是鸭嘴龙类的早期研究历史中,最具影响力的研究之一。鲁尔与莱特推翻之前的水生植物理论,认为鸭嘴龙类是水陆两栖的动物,而以陆地植物为食。鲁尔与莱特注意到鸭嘴龙类的复杂齿系,并将它们与马的牙齿相比较,发现两者有类似的排列方式,而且这些恐龙的牙齿会不断生长、替换。但他们认为,鸭嘴龙类的颌部不类似现代爬行动物,也没有马进化过程中所遭受的演化压力,所以无法解释这些齿系的功能。鲁尔与莱特认为禾本科(草)的份量太小,因此推测鸭嘴龙类不会以草为主食,而是以木贼为主食;在鸭嘴龙类化石的所处地区、地质年代,木贼是常见植物之一,而且富含淀粉、硅,在鸭嘴龙类的牙齿生长、替换过程中,硅可补充所需物质。鲁尔与莱特认为鸭嘴龙类的次要食物包含柔软的陆地植物、水生植物。鲁尔与莱特认为鸭嘴龙类的生活方式,类似现代的驼鹿,以陆地、湿地的植物为食。他们认为鸭嘴龙类的口鼻部、鼻管结构复杂,使它们可以将嘴浸泡在水中,以水中植物为食,如同现代驼鹿。
此外,鲁尔与莱特推测,鸭嘴龙类的嘴部两侧有肌肉组织,类似哺乳类的颊肌,而不像肉食性恐龙、现代爬行动物,口嘴部两侧没有肌肉组织。他们推测,当鸭嘴龙类进食时,会将植物置于两侧的颊部,使用颊齿磨碎植物。这个理论的根据是,鸭嘴龙类的上颌、下颌两侧,都有一道明显的棱脊,而上颌与下颌的牙齿位于内侧深处,显示在生前可能有肉质组织连接至上颌、下颌两侧。鲁尔与莱特也根据颌部关节,推测鸭嘴龙类的下颌只能做出单纯的上下动作,而无法做出前进、后退的动作。这种上下的动作,已足够将植物切成碎片。除了肉质颊部,鲁尔与莱特也推测鸭嘴龙类有相当发达的牙齿。
恐龙文艺复兴:陆栖鸭嘴龙类
在1960到1970年代,约翰·奥斯特伦姆(John Ostrom)提出许多新的恐龙生理学理论,造成小规模的科学革命,称为恐龙文艺复兴。在1964年,奥斯特伦姆曾发表一篇鸭嘴龙类研究,认为鸭嘴龙类既不是两栖动物,也不会以水生植物为食。奥斯特伦姆推测,鸭嘴龙类是完全的陆栖动物,以陆地植物为食。他根据鸭嘴龙类的坚牢齿系,推测它们能够以质地坚韧的植物为食,例如:富含硅、多纤维、木本的植物。奥斯特伦姆推翻鲁尔与莱特的进食方式理论,认为鸭嘴龙类的下颌关节可以做出上下、前进后退的复杂动作。但奥斯特伦姆不对颊部理论发表意见。奥斯特伦姆详细地推测鸭嘴龙类的进食方式,它们会将嘴部凑近树枝,用喙状嘴咬下树枝、树叶,在以头部力量向后拔。在1922年发现的胃内含物,可证实奥斯特伦姆的假设。鸭嘴龙类的其他生理特征,也证明它们是陆栖动物。它们具有骨化肌腱,沿者脊椎的神经棘排列,如果鸭嘴龙类在水中游泳、生存,这将妨碍它们的动作。奥斯特伦姆也提出,在鸭嘴龙类化石的所属地层,很少发现水生植物的花粉化石,显示鸭嘴龙类的生存地区、年代,水生植物并不常见。
近年理论在1984年,大卫·威显穆沛(David B. Weishampel)提出新的相关研究。他研究鸟脚下目的头颅骨骨缝结构,发现这群动物有非常特殊、灵活的上下颌结构,也包含鸭嘴龙类在内。当它们的下颌往上闭合时,左右上颌会往两侧稍微撑开;同时,上下颌的牙齿会互相磨合,可以磨碎位于嘴部两侧的植物。在2008年,以Natalia Rybczynski为首的美国、加拿大科学家,使用电脑模型测验威显穆沛的理论,认为鸭嘴龙类不可能做出这种动作。而在2009年7月份的《科学》,另一群科学家则做出不同结果的研究,支持鸭嘴龙类可做出这种动作的理论。
在2008年,科罗拉多大学博尔德分校的学生Justin S. Tweet研究一个短冠龙的标本时,在胃部区域发现一团细微树叶碎片[25][26]。Justin S. Tweet研究过这团细微树叶碎片后,认为这个动物是以较高植被为食,而非低矮的植被。对此,Mark Purnell表示胃部的食团不能准确地代表该动物的日常主食。
另外,根据某些白垩纪晚期鸭嘴龙类的粪化石,显示它们有时以腐烂的树木为食。树木并非高营养的食物,但分解的树木带有真菌、以生物碎屑为食的无脊椎动物,却是高营养价值的食物。
2009年:牙齿磨损痕迹研究,在2009年,英国莱斯特大学的Mark Purnell、Vincent Williams、以及伦敦英国自然历史博物馆的保罗·巴雷特(Paul Barrett)共同研究鸭嘴龙类的咀嚼方式与食性。他们决定采用新的研究方式,研究牙齿磨损痕迹的大小、方向、形态,试图归纳出鸭嘴龙类的嘴部运动方向,并进一步推测它们的食物的质地与结构。
巴雷特、Purnell、Williams以埃德蒙顿龙作为研究对象,埃德蒙顿龙的化石来自于美国、加拿大的白垩纪晚期地层,他们研究美国的埃德蒙顿龙化石,分析数百颗牙齿的磨损痕迹。他们仔细地清洁、处理颌部与牙齿化石,并将牙齿镀上一层金,以更清楚地辨识出牙齿的细微表面结构,并使用高分辨率的电子显微镜,以各种角度拍摄每颗牙齿,最后建立出牙齿的3D立体模型。这项研究发表于2009年6月份的《美国国家科学院院刊》,是第一个关于恐龙牙齿磨损痕迹的定量分析。
巴雷特等人发现鸭嘴龙类可能发展出不同于任何现代草食性动物的进食模式。借由比对每颗牙齿的3D立体模型,他们发现可以归纳出四种不同方向的牙齿磨损痕迹,而每种方向的磨损痕迹,可能代表特定方向的嘴部运动。根据这个理论,鸭嘴龙类的嘴部运动,共有上下、前后、以及侧向等不同方式。与现代草食性哺乳动物相比,现代草食性哺乳动物是利用灵活的下颌关节,做出复杂的前后、左右动作,以咀嚼嘴中的食物;而鸭嘴龙类则是借由上颌骨头的关节。当鸭嘴龙类的上下颌闭合时,上颌的左右骨头会向两侧撑开,而下颌牙齿以较内侧的位置,与上颌牙齿磨合。借由反复的上下颌闭合运动,嘴部两侧的食物会逐渐磨碎。除了上下颌的闭合动作,巴雷特等人也假设,鸭嘴龙类可以做出类似咀嚼的细微动作。
在同一份研究中,巴雷特等人也提出鸭嘴龙类以哪些植物为食的假设。但他们也提出警告,与鸭嘴龙类的嘴部运动理论相比,鸭嘴龙类的食性理论,有较多的部分是根据假设。巴雷特等人发现,这些牙齿的磨损痕迹相当平均,显示鸭嘴龙类在进食时,是重复地做出类似的嘴部运作。巴雷特等人假设,鸭嘴龙类可能主要以较柔软的树叶为食,而非较坚硬的树枝、茎;因为鸭嘴龙类必须采用特定方向、较大力的嘴部运动,才能咬断较坚硬的树枝、茎,这样会产生不平均的牙齿磨损痕迹。巴雷特等人也发现,这些牙齿没有明显凹陷痕迹,而以此推论鸭嘴龙类的食物中,带有微小的砂砾,或者是硅微粒。由于以低矮植物为食时,会吞入较多的微小砂砾,而禾本科(草)会带有较多的硅微粒。两种可能都显示,鸭嘴龙类可能以低矮植物为食,而非较高处的树枝、树叶。在白垩纪晚期,禾本科已经出现,但不是繁盛的植物,因此巴雷特等人推测禾本科不是鸭嘴龙类的主要食物来源。他们推测鸭嘴龙类的主要食物是木贼,木贼是白垩纪晚期的繁盛植物之一,高度低矮,富含硅。
巴雷特等人宣称,这项定量分析研究以牙齿磨损痕迹作为研究对象,不仅提出鸭嘴龙类的进食方式、主要食物的理论,也提供新的研究动物进食方式、食性的研究方法,未来也可以应用在已灭绝动物的研究上,例如恐龙、史前鱼类、与史前哺乳动物。俄亥俄大学的古动物学家劳伦斯·威特默(Lawrence Witmer),对这项研究成果表示高度赞扬,但仍质疑鸭嘴龙类在进食时,上颌是否会向外撑开的理论。
本词条内容贡献者为:
李学强 - 教授 - 宁夏大学

 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国