

某些真菌,细菌,绿藻和原生动物进行有性生殖时,两个细胞互相靠拢形成接合部位,并发生细胞壁融合而生成接合子,由接合子发育成新个体,这样的生殖方式称为接合生殖。接合生殖的两个个体,供体即雄体的部分染色体可以转移到受体(雌体)的细胞中并导致基因重组。水绵的有性生殖、草履虫的有性生殖都是接合生殖的典型例子。1
如水绵接合生殖时,阳性接合细胞内全部原生质通过接合管到达阴性接合细胞,由两个细胞的原生质融合而生成接合子;接合子经过减数分裂生成有性孢子各2个,有性孢子萌发为水绵的营养体。
释义接合生殖接合生殖是有性生殖的低级阶段,在低等生物类群中多发生这种生殖方式。接合生殖是由两个亲体细胞相互交换一部分核物质后,再分开进行分裂生殖的一种有性生殖方式。2
形态分类原生动物的接合生殖多见于纤毛虫类,按接合的双方,即接合子的形态又可分为两类:
①同配接合:接合子的形态相同。接合时双方暂时融合,小核在减数分裂后进行交换,相互受精后分开,如尾草履虫。接合双方紧靠在一起,口部融合,然后大核消失小核分裂两次,成4个,其中3个退化,一个再经一次分裂成为一个动核和一个静核。此时接合个体互相交换动核,然后静核与换来的动核融合。接着虫体分开,每个个体的融合核分裂三次形成8个核,其中4个成大核,3个退化。此后大核不分裂,剩下的小核与虫体同时分裂两次而成为4个新的子体。3
②异配接合:见于原毛目类纤毛虫。在进行接合生殖前,虫体先经一次不均等分裂,除小核外大核和虫体都分成大小两部分,成为大接合子和小接合子,前者固着,后者为自由游泳。小接合子找到大接合子后即牢固附着在其上开始接合。在接合过程中,合子核只在大接合子中形成,小接合子为大接合子吸收,其中钟虫就是一个典型的例子。3
接合类型根据接合管产生的位置的不同,接合生殖又可分为梯形接合、侧面接合等类型。4
水绵的有性生殖为接合生殖。其中又包括梯形接合、侧面和直接侧面接合3种类型,但以梯形接合为最常见。5
相关实例大肠杆菌大肠杆菌有两种性别系,当不同性系的个体杂交时,阳性菌体细胞接触阴性菌体细胞,两个细胞侧面形成接合管,阳性系细胞的DNA通过接合管流入阴性系细胞内,形成接合子;接合子经过细胞分裂形成两个大肠杆菌。
水绵水绵接合生殖多于春季和秋季发生,这时丝状体的颜色也由绿 变为黄绿。水绵接合生殖过程是:首先为2条水绵丝状体并行靠近,在两条丝状体的细胞相对的一侧各产生1个突起。继而两相对突起伸长,顶端接触,端壁融解形成1条连通的管子,叫做接合管。同时,各个细胞内的原生质体浓缩,形成1个配子。其中1条丝状体的每个细胞中的配子,以变形虫式运动通过接合管移入另1条相对细胞中,与其中的1个配子融合,产生1个卵形或椭圆形的合子,也称接合孢子。这样,1条丝状体的细胞都变空,另1条丝状体的细胞中都各有1个合子。空的丝状体可视为雄性,有合子的丝状体可视为雌性。以上生殖过程就叫做接合生殖。由于在两条丝状体之间有很多横向的接合管,外观上看颇像个梯子,故称做梯形接合。以后合子分泌产生厚壁,藻体死亡崩解后,合子沉入水底休眠。条件适宜时萌发,合子核首先经过减数分裂,所产生的3个单倍体核退化,仅1个发育,最后产生1条新的水绵丝状体。5
变为黄绿。水绵接合生殖过程是:首先为2条水绵丝状体并行靠近,在两条丝状体的细胞相对的一侧各产生1个突起。继而两相对突起伸长,顶端接触,端壁融解形成1条连通的管子,叫做接合管。同时,各个细胞内的原生质体浓缩,形成1个配子。其中1条丝状体的每个细胞中的配子,以变形虫式运动通过接合管移入另1条相对细胞中,与其中的1个配子融合,产生1个卵形或椭圆形的合子,也称接合孢子。这样,1条丝状体的细胞都变空,另1条丝状体的细胞中都各有1个合子。空的丝状体可视为雄性,有合子的丝状体可视为雌性。以上生殖过程就叫做接合生殖。由于在两条丝状体之间有很多横向的接合管,外观上看颇像个梯子,故称做梯形接合。以后合子分泌产生厚壁,藻体死亡崩解后,合子沉入水底休眠。条件适宜时萌发,合子核首先经过减数分裂,所产生的3个单倍体核退化,仅1个发育,最后产生1条新的水绵丝状体。5
草履虫草履虫接合生殖时,每个虫体大核消失,小核减数分裂生成4个核,其中3个核消失,留下的一个核分成动核和静核;动核通过接合膜交换,分别与对方的静核融合;接合结束后,两个虫体分开,各自经历三次核分裂和两次胞质分裂,形成4个新个体。
(一)草履虫的接合型
草履虫种内存在不同的繁殖群(syngen),每一繁殖群内存在不同的接合型(matingtype),只有同繁殖群内不同的接合型之间才能发生接合,进行接合生殖。在不同繁殖群之间,不同的接合型间一般不发生接合。6
有的草履虫根据繁殖群之间的这种有性生殖的隔离现象,这些繁殖群已被上升为不同的姊妹种,如原来的双小核草履虫(parameciumaurelia)的1个种已分成14个姊妹种(siblingspecies)。其它草履虫仍保留繁殖群(syngen)这一种下的分类单位。根据草履虫种类的不同,每一姊妹种或繁殖群内接合型的组成也不同,如双小核算履虫((P.aurelia)中的每一姊妹种和尾草履虫(P.caudatum)及多小核草履虫(P.multmicronucleatum)中的每一繁殖群内只有两个可发生接合的接合型,而绿草履虫(P.bursaria)和旋毛草履虫(P.trichium)的每一繁殖群内则有多个接合型,其中任何两型之间都可接合。因此接合型不同于其它动物的“性”。因为“性”不可能有多个,又因草履虫接合时每一个体内都产生两种原核,相当于精核和卵核,接合时精核互换与对方的卵核受精,故草履虫属于雌雄同体的动物,个体之间无性别之分。草履虫的不同接合型属于不同无性系(clone),无性系追溯到它的起源也可称之为大核系(caryonide),因为草履虫的无性繁殖主要是靠大核的不间断的分裂。
草履虫接合生殖时由小核产生受精作用重建新的大核,不同草履虫接合一次建成的新大核数目不同,一般每一接合后体不是建成4个就是建成2个,接合后体通过整个细胞的分裂各分得单一的新大核,从此由新大核通过无性分裂传下的后代群就叫大核系,亦即无性系。草履虫的不同接合型就分布在不同的大核系也即无性系中。6
(二)获得草履虫接合型的方法
获得草履虫纯的不同接合型有多种方法,这里介绍无性系法。
任何草履虫在它的繁殖地都至少存在一个繁殖群内的两个或多个接合型,它们由不同的无性系来体现。将由自然环境中采得的草履虫单个分离,用原地过滤并煮沸放冷的池水洗去一切其它生物,将这些草履虫放入凹玻片的凹坑内,加入稻草汤培养液(制法见后),放入20℃-25℃下培养,数日后即繁殖成群,再移入试管培养。虫体增多后要每日至少加一次新培养液,以保持其旺盛的无性繁殖。这样建起的多个无性系,必然包含相对的接合型。在适宜条件下,将这些无性系中的每两个取样相混,凡能发生接合的两个无性系就是两个相对接合型。找出相对接合型后,保持其食物充分和生长旺盛,就随时可取样相混,即可获得接合对。7
如欲保存以后备用,可放入4℃-6℃冰箱中,每半月到1月加一次新培养液,可长期保存接合型。用时升温加食,经过几天快速繁殖后,相混仍可产生接合反应。
(三)控制草履虫接合的方法
草履虫能否进行接合生殖取决于内在因素和外在因素,即接合型、成熟度、营养状况、培养液的pH值、培养温度和虫体密度。因此,草履虫的接合生殖是可以诱导的。6
如前所述,革履虫种内或繁殖群内存在不同的接合型,只有相对接合型之间才能有接合反应,进行有性生殖。属同一接合型的不同无性系之间一般不能接合。
草履虫的一个大核系的一生可分为四个阶段,即未成熟期、成熟期、衰老期和死亡期。未成熟期是指新生的大核系只经较少次数的无性分裂阶段。处于此期的草履虫即使是属于相对的两个接合型,相混后也不会有接合反应,即使有百分率也非常低。只有达到成熟以后,相对接合型之间才可能出现接合反应,而且有较高的接合率。处于衰老期的虫体由于代谢及生理上的原因,虽属相对接合型的个体相遇,接合率也会明显降低,而且接合后体的成活率也显著下降。
达到成熟期的相对接合型能否相混后立即接合还取决于虫体的营养状况。过度饱食或过度饥饿都不能出现接合反应。因此,想得到理想的接合材料必须控制食物。只有旺盛繁殖后,由饱食转入饥饿的数小时内,两型相混才会立即发生接合。发生接合的温度在15℃-30℃之间,20℃-25℃为最佳。上述四个因素是控制接合不可忽视的。
草履虫培养液可按下法制出:取10克稻草或麦杆加水1000毫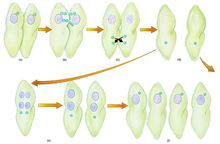 升,煮沸过滤,放凉后接种产气杆菌属(Aerobacter)的细菌,放入30℃下培养3天,稻草汤变混浊后即可使用(如无产气杆菌,也可在空气下暴露3天后使用)。将已建起的无性系用此培养液培养数日,高速繁殖后都可达到成熟期,当加食后待培养物开始变为清彻透明时,即将达到饱食转入饥饿的时期,这时将属于相对接合型的两个培养物相混,立即或在数小时内发生接合。接合初期的反应是几个甚至几十个虫体相互粘着、聚团。过1小时以后将出现大量的接合对,开始接合生殖的内部过程。用这些方法既可找出相对的接合型,也可难持接合型,随时相混即能获得供实验用的接合材料。
升,煮沸过滤,放凉后接种产气杆菌属(Aerobacter)的细菌,放入30℃下培养3天,稻草汤变混浊后即可使用(如无产气杆菌,也可在空气下暴露3天后使用)。将已建起的无性系用此培养液培养数日,高速繁殖后都可达到成熟期,当加食后待培养物开始变为清彻透明时,即将达到饱食转入饥饿的时期,这时将属于相对接合型的两个培养物相混,立即或在数小时内发生接合。接合初期的反应是几个甚至几十个虫体相互粘着、聚团。过1小时以后将出现大量的接合对,开始接合生殖的内部过程。用这些方法既可找出相对的接合型,也可难持接合型,随时相混即能获得供实验用的接合材料。
(四)观察草履虫接合过程的方法
由上述方法获得理想的接合材料,用定期压片(根据情况每隔10-30分钟),石炭酸品红染色的方法做临时装片,这样就可以观察到接合过程中整个核的变化情况。
具体方法如下:
用离心机的最低转数将接合材料浓缩,吸1滴置于载玻片的中央或者用微吸管吸几个接合对于载玻片中央(切记水不要过多),在虫体周围点4点浆糊,然后加上盖片,用镊子或铅笔尖在盖片的四角向下压直至虫体只能缓慢移动为止。
在盖片的一侧滴加石炭酸品红染液,在对侧用吸水纸吸去多余的染液,染15分钟后即可观察。这样的临时装片也可经梯度酒精脱水,二甲苯透明,树胶封片,做成永久装片。盖片在脱水过程中脱落,可将载片和盖片分别封成两张片子,上面都会留有接合的虫体。
本词条内容贡献者为:
赵磊 - 副教授 - 中国农业大学
 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国