

《卷首诗》
我们是谁?我们从哪里来?要到哪里去?
这是一个古老而又永恒的话题。
迄今为止,在人类所能观测到的数百亿光年的宇宙范围内,
地球是唯一的生命绿洲。
这个小小的绿色星球,如同一座富饶的岛屿,
拥有制造生命的神奇魔法,在空旷寂寥的宇宙中,
孤独而又自豪地高唱着生命之歌。
因为绝无仅有,所以弥足珍贵。
生命的诞生是机缘巧合还是预谋已久?
那个驱动有机物命运的齿轮如何开始转动?
是大洋深处最初的一丝蠕动,还是池塘河边露出的一线生机?
生命起源问题,和黑洞、暗物质、暗能量以及宇宙起源、意识起源,
并称为“一黑两暗三起源”,向来是科学问题上的明珠!
这是个发生在近40亿年前,绝大部分物质证据都早已湮没的问题。又是一个我们已知的世界中只发生过一次的孤立事件,
没有可以归纳的基础。
同时它横跨生物、化学、地理、地质、气候、海洋、宇宙学诸领域,涉及的已知和未知千头万绪。
最麻烦的是,即使有人真的发展出和事实相符合的理论,
也几乎没有可能实地验证—这个实验本身就是个可能耗时千万年,
或者上亿年的局中局。
然而这丝毫没有驱散各领域科学家几乎是无休无止的兴趣。
自从宗教的枷锁打破之后,科学性的生命起源假说已不下几十种,
争论不休,聚讼纷纭,辨伪存真,谁借慧眼?
自然发生说、有生源论、生命永恒论、宇宙胚种论、化学进化论......
亿万斯年,光的世界,大道至简,万法归一。
鸿蒙开天辟地,生命缘何而起?你问我,我问谁?
是“宠辱不惊,闲看庭前花开花落;去留无意,静观天边云卷云舒”,
还是“凭栏观日月,无语问苍穹”?
是“对长亭晚,骤雨初歇......执手相看泪眼,竟无语凝噎 ”,
还是“国难思良将,家贫思贤妻,有事钟无艳,无事夏迎春”?
“一生经过彷徨的挣扎,自信可改变未来,问谁又能做到”
这首歌致敬了曼德拉;
“国有疑难可问谁,强国一代有我在!”
这句话来源于国科大。
只有在科学昌明、创新技术发展到今天:
在显微镜开启了微观生命科学探索 ,
X射线开启了生命医学影像探索,
量子力学推动了宏观生命科学探索的时代,
我们才稍具条件发起对这个问题的攻关。
科学突飞猛进,科技自立自强;
随着各学科的进步不断演进,不断丰富细节和淘汰伪说。
本书汇聚涓滴力量,引领你走进地球原初不毛之地,
破解生命起源哑谜,揭示万千生灵进化的非凡之旅。
有诗为证:
千流归大渊,万马踏平川。
聚澜腾细浪,高路入云端。
第一部分 为什么总是讲不清“生命起源”问题:方法与意义
I
生命几乎已经征服了地球上的每寸土地。但在地球刚刚形成时,它实际上就是个没有任何生机的石球。那么生命是如何在这个星球上诞生的?生命从何而来?又将走向哪里?它们如何从无到有,从简单到复杂的?最初是什么样子的?这些问题一直矗立在人类最为着迷和困惑同时又尚未解决的科学难题中,或多或少扰动过你的心灵。
这个可大可小的问题,少数人----科学家、哲学家或者只是一个有抬头仰望星空、低头关心根源的普通人,视之为最重要的事绞尽脑汁去思考,甚至耗尽毕生也不放弃;多数人则毫不触及或者忙于俗世生活避而不谈。他们未来的大脑里,没有这么宏大问题;所以理所当然丢给哲学家们去思索了,或者满不在乎的任神学家手握答案。
而于我则认为,思考这些问题的答案,对于理解我们自身以及我们所居住的这个世界,都具有无法估量的价值。因为真正了解过去才能明白未来,这不仅是对过去的探寻,也是对生命本质的深刻思考。对于个体而言,或可以唤起童年时的好奇心而激发思考的火花,或可以让人在难题面前始终保持谦逊的求知态度、敬畏大自然,而拥有这种好奇心和对真理不懈追求的愿景,对科研人员或者青少年而言,无疑是很有益处的。
达尔文曾断言:“目前关于生命起源的思考纯粹是废话”,现在已经不再成立。达尔文所处的时代连基因是何物都还不知道,现在已经是20世纪20年代,三域生物所有RNA聚合酶的结构类型均被阐明,“最后一块拼图”也终于被中国科学家补上。
但是,正如《生命起源的奥秘:再评目前各家理论》指出:“我们在合成氨基酸方面的成就有目共睹,但合成蛋白质和DNA却始终失败;两者形成了强烈的对照”。虽然,科学发展到今天,我们能以极大地效率在实验室利用机器合成出需要的生物大分子,在生命起源前环境里的合成实验却“始终失败”,这或许是大自然跟人类开的一个玩笑吧,以此提醒人们面对大自然、面对时间长河应当保持谦逊的姿态。
II
西方人早期谈论生命起源问题的思路可以分为两类:
要么是拿出基督教的《创世纪》,认为“生命”起源于空白和混沌,然后认为是由“神”创造了“秩序”,至于什么样的秩序,就是“人类”的事;
要么在早期的生命发生学中,大多数人认为生命是自然发生,本质是突显论或自然发生论,如青蛙可以从湿土中产生,老鼠可以从腐烂物质中产生,昆虫从露水中产生,蛆从烂肉中产生等。温暖、潮湿、阳光乃至星光都常被说成是促进生命自然发生的有益因素。包括像古希腊的亚里士多德,甚至是近现代的诸多哲学家、科学家,如哥白尼、培根、伽俐略、哈维、笛卡儿、歌德和谢林都不自觉的接受了“突显论”的思想。
在《物种起源》出版之前,科学家公认的模式是培根(Francis Bacon)的归纳模式,即在实验的基础上大量采集资料,然后对资料进行总结归纳,得出一个简洁的结论,也就是新的知识。达尔文应用的是假说演绎法,先立假说,再去演绎,推出一些新的认识,然后去自然界寻找证据来证明这些认识,这样就验证了假说。
恩格斯在《路德维希·费尔巴哈和德国古典哲学的终结》的第二章,是这样描述的:哲学家依照他们如何回答这个问题而分成了两大阵营。凡是断定精神对自然界来说是本原的,从而归根到底承认某种创世说的人(而创世说在哲学家那里,例如在黑格尔那里,往往比在基督教那里还要繁杂和荒唐得多),组成唯心主义阵营。凡是认为自然界是本原的,则属于唯物主义的各种学派。明显,神创与自然发生已经归于陈旧哲学。
而在近现代,则出现了一系列新的生命起源假说。在陈阅增第四版的《普通生物学》中则写了两类比较有代表性的有关原始生命的假说:
其一是“蛋白质起源说”:即“奥巴林”(亚历山大·伊万诺维奇·奥巴林(Oparin,Alexander Ivanovich))和“福克斯”分别提出“团聚体学说”和“微球体学说”。奥巴林将多肽、核酸、多糖、磷脂的溶液摇晃混合后,发现在胶体溶液中的大分子凝聚形成直径约1-500微米的团聚体小滴。这个团聚体小滴在奥巴林看来是有代谢的特性,因为其能从周围介质中吸取不同的物质,然后还出芽分裂成效的团聚体小滴,所以,奥巴林认为这就是一种生命起源解释方式。
但这显然有很大问题!因为这极大可能是一种物理或化学特性,而绝非代谢,因为代谢指向的是具有“供能”性质的糖类、脂肪等,你绝不能说它是生命。而“福克斯”的微球体学说,则是将“类蛋白”和“核酸”加热浓缩,最终形成了直径约为1-2微米的胶质小体,有双层膜,这类微球体可以选择性吸收胶质中的类蛋白而生长和繁殖,这就体现出某种生命特征,但这其实和奥巴林的团聚体学说本质没有太大差别;
其二是“核酸起源说”:此理论认为“RNA”是地球上最先出现的第一批基因和酶,并不是DNA和蛋白质,而且推测混合的核苷酸单体可“自发”结合成短链RNA。短链RNA作为第一基因以自身编码的信息为模板,短链RNA又自带“催化性生物机能”进行自我复制,即RNA是第一个作为具有“催化性生物机能”和能作为基因载体的分子,被称为“RNA世界假说”。对于核酸起源说,认为RNA既能作为基因载体,又能作为酶,这显然在某种程度上是有道理的,说是最早的生命也可以,但最早的RNA是如何一步一步发展而来?
所以,在陈阅增第三版的《普通生物学》中,“生命起源”其实并没有完全说清楚,更多是拿来主义的借用了西人的研究成果。而要真正说清生命的起源必须要说清分子的演进过程,以及生物演化的中轴,毕竟“生命起源”是夹在两者之间,没有横跨分子与生物的底层原理,就不可能真正说清楚“生命的起源”。
III
【近期,美国斯克利普斯研究所证明了二酰胺磷酸盐(DAP),可通过化学反应将脱氧核糖核苷构建编织成原始DNA链。相关研究成果发表于《德国应用化学》。】
这一发现指出“DNA和RNA”可能是类似化学反应的共同产物。即是说,第一批自我复制的分子作为地球上第一种生命形式,可能是两者的混合物。而质疑“RNA世界”假说,部分原因是RNA分子太“黏”,它擅长吸引并黏附其他单独的RNA结构单元,形成镜像链,这个新链的每个结构单元都与原始模板链的结构单元互补。如果新链可以脱离模板链,并通过相同的过程开始模板化其他新链,那么它就可以实现构成生命的自我复制。但它们却不擅长与这些链分离。所以,在早期无解链酶的世界里,是很难让RNA链分开的,证明RNA世界假说本身也不一定绝对正确。
然后是,在罗斯((英)罗斯(Rose, S.))·著的《大脑的未来》中写道:“生命是涉及碳、氢、氧、氮构成的化合物之间的相互转化和相互作用,以及和钠、钾、氯、硫、磷等离子和重金属等反应,然后基于原始地球充满氮气、甲烷和二氧化碳更适合还原而非氧化的环境,以及伴随原始地球猛烈的雷鸣电闪等恶劣环境,最终使得这些基本的元素化合物逐渐生成“有机分子”,从糖类、氨基酸、核苷酸到蛋白质、脂肪、RNA、DNA等生物大分子的有机物稀汤。”
而后,“罗斯”直接就认为生命的开始居然是源于脂或油,是油滴中聚集了有机物以及钠离子和钾离子,从而构成最原始的原核细胞。这种说法其本质就是认为“初始细胞”源于“随机组合”,这就相当于一阵大风,将废品站的铜铁铝吹成了现在登机站台下的一架驾簇新的飞机,或者让一只猴子学会盲打之后,打出了莎士比亚的《罗密欧与朱丽叶》,生命不可能有这样环境去大把大把的浪费时机。
IV
科学研究的一般规律是,把自然规则研究清楚透彻,然后应用到实验室生产出人类的福利,即从大自然到实验室,一种对规律的应用;而在用进化理论试图解决“起源”的问题上,既然是从无到有,实验室就不能借用已找到的规律,而是要生产出一套规律去套用到大自然上,这是跟通常科学研究的方法是不同的,这是还原论的困境。
科学性的生命起源假说有不下几十种,随着各学科的进步不断演进,不断丰富细节和淘汰伪说,但是归根结底从方法上来说只分为两类,一类是正向的还原法,即不断搜集地球生命初始时代遗留下来的蛛丝马迹,包括化石、碳痕迹、同位素异常等等,不断试图还原当时的大气、海洋或者地质化学条件,试图通过模拟的实验,一点点验证生命起源的历程。这是传统的方法,它搜集和整理了大量的证据和数据,尤其在年代测定方面发展了很多高明的技术,取得了很多成果。到目前为止,最早的生命活动残迹可以上溯到38亿年之前,是由极其古老的岩石中碳同位素扰动间接证明的(生命是已知唯一改变碳同位素比例的自然过程)。
另一类是反向的反演法。最早的生命虽然早已死亡,但是它们的后代充满地球生物圈。这些后代种类极其繁杂,但是它们在分子生物和生物化学的层面具有令人惊奇的共性。这除了让科学研究者更加坚定了“生命同源”的信心之外,还提供了一个新的思路:共性即共祖特征。共性越多的生物,它们的亲缘关系越近。越普遍的共性,越有可能是生命在遗传树“根部”的特性。绝对普遍的生物共性,几乎一定是生物起源时的特性。跟传统的还原法正好相反(还原法是苦于证据太少太不完整),现代的生理学和基因科学为反演法提供了海量的各种生物遗传信息。整个生物圈的数码化遗传信息:基因组都是证据,只看研究者如何去挖掘和解读。难在如何穿透近40亿年生物分化、变异和灭绝造成的数据迷雾。如果我们能反推出最初的生物是什么样的特性,则如何起源的问题就解决了大半。因为在生物学中,“你是什么”基本就表述了“环境怎样造就了你”。反演法的研究最喜欢的一个术语就是LUCA(Last Universal Common Ancestor):“最后的共同祖先”。它是一个定义在逻辑上的生物,是现存所有生物的共同祖先。(之所以说最后,是因为不排除它之前更早的分支已灭绝且未留下后代)
虽然生命起源问题是个难以索解的问题,没有人能够目睹生命形成的过程,科学家研究的成果应该说都还只是假说、理论,不是绝对的事实,正如霍金提出的宇宙大爆炸理论(Big Bang theory),不管科学怎么进步,如果这一理论是正确的,它本身意味着我们只能知道大爆炸发生以后的事情,大爆炸之前的情况只能是推理和想象,猜测便指明了无法得到百分之百的证明(不管有没有事物存在或存在什么事物)。
V
但我们终将不该畏难,不能畏而不前。人类的创造力是无穷无尽的,以无穷无尽的创造力面对无穷无尽的科学和艺术难题,我们会创造出崭新的思考框架和理论逻辑。我们不断扩大这研究的“方法库”,这些方法来源却不限于:
考古生物学与生命起源的化石证据;
在南非的巴伯顿绿岩带,科学家们发现了约34.2亿年前的微化石,这些微化石是由特殊类型的甲烷循环微生物组成的,这是迄今为止发现的最古老的生物化石之一。这些微化石的发现不仅为我们提供了了解生命最初是如何开始的线索,也表明生命起源的时间非常古老,早于地球上的许多其他生命形式。
另一方面,对现代生命形式的基因和细胞结构的研究也为我们提供了关于生命起源的重要线索。例如,科学家们通过对细胞的研究发现,细胞核是真核生物细胞中的重要结构之一,它可以储存遗传信息并指导蛋白质的合成。而这种细胞结构的出现,可以追溯到大约20亿年前的一种原始真核生物。
此外,生物分子的化学结构也是支持生命起源理论的重要证据之一。例如,核酸和蛋白质是构成生命的基本分子,它们具有复杂的化学结构和功能。通过对这些分子的化学结构进行研究,我们可以了解它们的演化历程和可能的起源。例如,研究表明,核酸和蛋白质可能是从更简单的化合物中演化而来的,这些化合物可能在早期地球的化学环境中形成。
b.实验室模拟生命起源的化学过程;
实验室模拟生命起源的化学过程可以被理解为一种在实验室环境中模拟和重现早期地球条件下生命起源的化学演化过程。这个过程主要基于对原始地球大气和环境的模拟,并通过实验验证生命从无机小分子物质到有机小分子物质,再进一步到更复杂的有机分子物质的演化过程。
这个模拟过程主要包括以下几个步骤:
创建模拟原始地球的条件:在实验室中,科学家们会模拟早期地球的条件,包括温度、压力、酸碱度、水含量等。此外,他们还会模拟原始地球的“还原性大气”,如氢气、氨气、甲烷和水蒸汽等。
模拟地球的化学反应:在模拟的原始地球条件下,科学家们会开始模拟地球早期的化学反应过程。这些反应包括但不限于水合、脱氢、加氢、脱氧、加氧等。
检测和记录反应产物:通过特定的检测手段,如光谱分析和色谱分析等,科学家们可以检测反应后的物质,并记录下它们的种类和数量。
分析并解释结果:根据检测结果,科学家们可以分析并解释这些产物是如何形成的,以及它们与原始地球条件的关系。
在这个过程中,科学家们的主要目标是理解早期地球的化学环境如何促进从无机小分子到有机小分子,再到更复杂的有机分子物质的转变。此外,他们还试图理解这些早期反应是如何影响地球的生命起源和演化的。
实验室模拟生命起源的化学过程是一个高度专业化的研究领域,需要专门的知识和技能。尽管这个领域的研究仍然有许多未解的问题,但它已经为我们提供了许多关于地球生命起源的新思考与研究。
c.计算机模拟、仿真与生命起源的数据;
计算机模拟在生命科学研究中的应用与进展主要表现在以下几个方面:
分子模拟与生物大分子:利用计算机模拟技术,科学家们可以研究生物大分子的结构和行为,如蛋白质和核酸等。这种技术可以帮助科学家们理解生物大分子的稳定性和动态性,从而加深对生物学过程的理解。例如,分子动力学模拟技术可以模拟蛋白质在不同环境下的构象变化,探索其稳定性和功能。
生物模拟与仿真:计算机科学可以用于生物模拟与仿真,即借助计算机模拟和重现生物系统的行为和特征。科学家们可以使用计算机模拟来研究生物系统的复杂性和动态性,从而更好地理解生物学过程。生物仿真则是指将生物系统的模型嵌入到计算机程序中,通过模拟真实生物系统的行为,帮助科学家们预测和研究各种生物过程和现象,例如药物作用、疾病模拟等。
生物数据分析:随着生物学研究的不断发展,产生的生物数据量也越来越大。计算机科学在生命科学中的另一个重要应用领域是生物数据分析。通过计算机算法和数据挖掘技术,科学家可以从大量的生物数据中提取有用的信息和模式,这些信息可以用于研究生物进化、疾病诊断和治疗等方面。
计算机科学的应用为生命科学研究提供了新的视角和方法,不仅加速了科学研究的进程,更为生物医学研究和应用提供了更多的可能性。
d.复杂性涌现与地球生命算法(后面章节会有涉及);
e.向不同学派与争议观点学习。
关于生命起源的其他学说还有化学起源论、异源传递论、智能设计论等等。
化学起源论:这个起源论指出生命最早的形式可能来自于地球上的化学反应和自组装,如蛋白质和核酸,由于它们能够在无机物条件下耐受变异和复制。这个观点是目前科学界较为接受的学说之一,但是科学家们仍在不断探索和验证这个观点。
异源传递论:这个起源论认为生命可能来自于宇宙中的其他星球,如随行的微生物或其他化学物质。这些微生物或化学物质通过陨石、彗星撞击等自然过程到达地球,并在适宜的条件下繁衍和演化。这个观点也是目前科学界正在研究的领域之一,但仍需要更多的证据来证明它的可行性。
智能设计论:这个起源论认为生命的起源来自于先有智慧的设计者。这种起源论通常被归类为“创造论”,其认为生命的存在不是自然演化或偶然性的结果,而是有意识、有目的地创造的。这个观点没有被广泛接受,主要是因为缺乏科学证据来支持它的正确性。总的来说,关于生命起源的研究仍在进行中,尚没有明确的答案。不同的学派和观点都有其优缺点和证据支持,科学家们仍在不断探索和验证这些观点,以更好地解释生命的起源和演化,我们可以在不同学派与争议观点学习,积累合理的思想成分。
f.还有一种比较另类的研究方法:LUCA模型的建立。这是从能量生物学的角度入手。因为与能量代谢相关的基因,正是所有生物中最古老,变异最慢,共性最大也最核心的基因。生物学家们讨论和试图描述LUCA的时候,不是关注它长什么模样,有多大个头,而是关注它执行何种氧化还原反应,有哪些代谢酶,生活在什么样的地理和化学环境。这已经完全是一种崭新的体系了。
VI
生命起源的意义及对人类意识的启示是广泛而深远的。
首先,生命起源本身是科学研究的一个重要领域。生命起源的奥秘是自然界的一大未解之谜,了解生命起源有助于我们更好地理解生命的本质和规律,为生物医学、遗传学、进化论等学科提供重要的理论基础。通过对地球最早生命形式的研究,我们不仅能更深入地理解自身的起源,也能更广泛地洞悉宇宙中生命的本质和可能性。当前,尽管科学家们已经取得了显著进展,如揭示了生命基本组成部分的合成途径,提出了RNA世界假说,以及在实验室里模拟早期地球环境,但生命起源的完整画卷仍有待进一步揭晓。
其次,生命起源对人类意识的形成和发展有着重要的启示作用。从生命起源的角度来看,人类意识并非是独立于物质世界的神秘现象,而是物质世界演化和发展的产物。在这个过程中,自然界的进化机制发挥了重要作用,而人类的意识也是这个机制的一部分。还有,地球外生命的可能性也是一个激动人心的研究领域,它不仅挑战着我们对生命定义的理解,也可能为我们提供关于生命多样性和适应性的新视角。
此外,生命起源的意义和目的对人类也有着重要的启示作用。人类作为自然界中的一员,需要认识到自身的价值和使命,同时也要意识到自己对自然界的影响和责任。在面对环境变化、生态破坏、气候变化等全球性问题时,人类需要承担起更多的责任和义务,未来的研究将继续探索这些问题,包括更深入地研究地球最古老的化石记录,进一步实验模拟早期地球环境,以及探索太阳系内外可能存在生命的环境,学习保护和改善地球生态系统,以确保人类和地球的可持续发展。
最后,生命起源的研究也为人类探索未知世界提供了重要的工具和方法。探索生命的起源,是一项挑战巨大但意义深远的科学征程。生命起源的研究充满了未解之谜。通过研究生命起源,我们可以更好地了解自然界的规律和机制,探索新的科学领域和未知的领域。特别是在分子生物学、地球科学和天文学等领域,通过探索和研究可以为人类提供新的技术和手段,推动科学的发展和进步。随着科技的飞速发展和进步,我们对生命起源之谜的理解必将更加深刻。
总之,生命起源的探索不仅是对过去的追溯,也是对未来的展望。它不断挑战着我们的知识疆域,引导我们在自然界和宇宙的广阔舞台上,继续寻找关于生命最根本问题的答案。
第二章 生命的地理起源(反推,七大可能地)
生命起源的问题,向来是生物学中的皇冠,同时也是Mission Impossible。这是一个发生在近40亿年前,绝大部分物质证据都早已湮没的问题。又是一个我们已知的世界中只发生过一次的孤立事件,没有可以归纳的基础。同时它横跨生物、化学、地理、地质、气候、海洋、宇宙学诸领域,涉及的已知和未知千头万绪。最麻烦的是,即使有人真的发展出和事实相符合的理论,也几乎没有可能验证—这个实验本身很可能耗时千万年,或者上亿年。然而这丝毫没有驱散各领域科学家对这个问题无休止的兴趣。
首先,这种兴趣被生命起源于哪里点燃。大约在40亿年前,地球上第一次出现了生命,它只是被包在某种囊中的遗传分子,长得跟我们当今所认识到的任何生物都不像,结构甚至比细菌的细胞还要简单。科学家一直在钻研这初始的生命究竟是如何形成的,不同的地点代表着不同的能量形式,不同的能量形式影响着不同的构造和场景,所以找到生命起源的地点也许是破解生命起源问题的关键。
我们总是设想,甲烷菌产生于甲烷多的地方,硝化细菌,产生于硝化物多的地方。硫化菌产生于硫磺多的地方,以此类推,不一而足,他们都有原产地,都有着土著特征。那么究竟在地球的哪个地方,藏这这样一个地方:它拥有能产生生命的关键元素,而且它的环境能供这些元素进行完美的化学反应。
 i小池塘猜想
i小池塘猜想
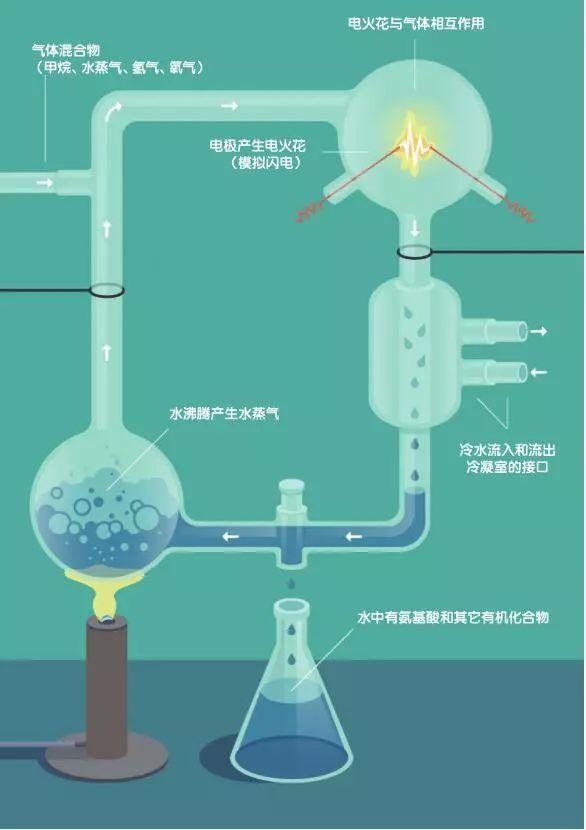
达尔文是最早开始尝试回答这个问题的人。1871年,他在一封信中描述了自己的猜想:有一个温暖的小池塘,里面富含化学物质和盐,环境中还有光、热和电。达尔文认 为在这样的环境中,蛋白质可能会自发地形成,然后会变成更复杂的有机体。20世纪50年代,美国化学家哈罗德·尤里和斯坦利·米勒进行了著名的米勒—尤里实验。他们建立了一个受控型密封系统,模拟地球早期大气层环境。
他们在长颈瓶中装上温水来模拟当时的海洋,当水蒸气蒸发时,会被收集在另一个烧瓶中。尤里和米勒在该实验装置中引入了氢气、甲烷和氨气,模拟早期大气层无氧气的状况。然后,他们释放电火花,来模拟闪电,进入这种混合气体构成的无氧大气层。最终,利用冷凝器将这些气体冷却成液体,收集进行分析。
实验结果表明,在冷却的液体中大量地存在着有机化合物,约有10%到15%的碳以有机化合物的形式存在。其中2%属于氨基酸,以甘氨酸最多。但核酸本身,如DNA或RNA则未出现。尤里和米勒得出结论称,有机分子形式能够来自于无氧大气层,同时最简单的生命体也可能孕育在这种早期环境中。
这场实验是20世纪最著名的实验之一,当时在社会上造成不小的影响,不过现在我们知道,当时在这个实验中生成的有机成分,并不足以构成生命,也意味着这场实验不足以说明达尔文的猜想是正确的。
生命想要“无中生有”,这三大要素必不可少:遗传密码、新陈代谢以及膜。遗传密码中携带可以用来制造细胞的生物蓝图;新陈代谢可以为细胞提供能量,而膜可以把这些成分和反应统统包裹在里面。在现存的有机生物中,以上三个要素都是由相同的原子构成的,即碳、氢、氧、氮、磷和硫。因此生命的摇篮至少要能提供充足的基本原子,还要有能让原子发生反应的基本条件。
ii海底起源猜想
 当生物化学家们还在苦苦思索这些问题时,深海探险家在太平洋地区有了惊人的发现。在1979年,美国有艘潜水艇潜入了距海平面2千米深的海底,在那里发现了海底黑烟囱,它的原理和火山类似,只不过是在海底喷发。在海底黑烟囱中,人们发现了不同寻常的完整的生态系统。于是科学家猜想也许生命起源于这些海洋底部的热泉。
当生物化学家们还在苦苦思索这些问题时,深海探险家在太平洋地区有了惊人的发现。在1979年,美国有艘潜水艇潜入了距海平面2千米深的海底,在那里发现了海底黑烟囱,它的原理和火山类似,只不过是在海底喷发。在海底黑烟囱中,人们发现了不同寻常的完整的生态系统。于是科学家猜想也许生命起源于这些海洋底部的热泉。
颇为巧合的是,大约在两年后,旅行者2号太空探测器发回了木卫二的照片,人们通过照片推测木卫二的冰壳下可能存在海洋。这个发现激起了人们对外星生命及其起源的兴趣,也给了相信海底起源说的人信心,甚至有人认为海底热泉既然能在地球上孕育生命,也一定能在其他星球起到相同的作用。
 但后来科学家们放弃了这个猜想,因为海底黑烟囱中存在的氢含量很低,而且它的温度太高,新生的分子难以在这样的高温中保持完整,如RNA中的遗传物质会在高温里被迅速降解。
但后来科学家们放弃了这个猜想,因为海底黑烟囱中存在的氢含量很低,而且它的温度太高,新生的分子难以在这样的高温中保持完整,如RNA中的遗传物质会在高温里被迅速降解。
21世纪之初,人们在大西洋中部发现了温度较低的海底喷口,这个地方喷发碱性的海底热泉。海水和海底矿物质在此处发生反应,生成带有微小孔隙的岩石,以及富含氢气的温暖液体。科学家认为岩石的气孔是早期生命出现的理想之处,尤其是碱性热泉和酸性海水之间存在电化学梯度,能自发形成乙酰磷酸和焦磷酸盐,这是两种能为活细胞供能的化学分子。它们也可以为分子合成提供能量,让溶解的二氧化碳和氢气合成有机分子,再构建蛋白质的组成部分和RNA。
iii 陆地起源猜想
如此看来,碱性的海底喷口似乎是生命起源的理想摇篮,只不过它也有自己的短板——过于潮湿。在地球上还没出现生命之前,生物分子是很罕见的,如果蛋白质的组成成分被过分稀释,那么它们就无法互相碰撞,最终形成蛋白质链。于是科学家猜想,或许存在一个有着干湿循环的地方,让最原始的分子能够碰撞出生命的火花,借此出现了陆地起源的猜想。
 陆地起源说认为,生命开始于有着周期性干涸的环境中,脱水作用在合成有机分子时扮演着重要的角色。每当一个蛋白质的组成成分被添加进蛋白质链中时,会释放出一个水分子。众所周知,酶是活细胞的催化剂。在生命的初期,酶还没有出现,只能依靠干燥的环境担任脱水的作用。
陆地起源说认为,生命开始于有着周期性干涸的环境中,脱水作用在合成有机分子时扮演着重要的角色。每当一个蛋白质的组成成分被添加进蛋白质链中时,会释放出一个水分子。众所周知,酶是活细胞的催化剂。在生命的初期,酶还没有出现,只能依靠干燥的环境担任脱水的作用。
 科学家们认为在火山岛上的淡水池比在深海喷口附近更容易形成脂肪膜。在海洋中,溶解的钙和镁离子会妨碍脂肪酸聚集在一起形成连续的膜;但是在淡水中,脂质可以轻易聚集,就像水油混合时,油会轻易和水分离,出现很明显的边界一样。
科学家们认为在火山岛上的淡水池比在深海喷口附近更容易形成脂肪膜。在海洋中,溶解的钙和镁离子会妨碍脂肪酸聚集在一起形成连续的膜;但是在淡水中,脂质可以轻易聚集,就像水油混合时,油会轻易和水分离,出现很明显的边界一样。
为了证明这个猜想,科研人员从美国黄石国家森林和加州拉森国家公园的温泉中采集样品,再让这些样品经历干湿循环。他们发现,如果他们将这些样品脱水然后再补充水分,其中类似RNA的分子就会被包裹在脂肪膜中,就像原始细胞膜能包裹遗传物质一样。
有的科研人员认为,比起火山水池,地热田似乎是更理想的环境,因为它更干燥。地热田是指用来释放地热蒸汽的热岩石。科研人员认为,这里的化学反应更接近于我们自己细胞内部的化学反应,因为地热蒸汽富含能孕育生命的重要元素。通过控制蒸汽浓度,小水滴的形成可以为生成RNA创造理想的环境。
要生成RNA,首先要制造核糖,也就是RNA中的R。创造R的原子可能早已存在在大气中,不过没有外力的帮助,这些原子不会自发变成我们需要的核糖。这时候硼酸盐矿物可以成为有效的助力,它能引导原子合成更多的核糖,而且还能巩固核糖的结构,以免它们分崩离析。因此,有科研人员认为沙漠才是生命的摇篮,因为岩石中有着丰富的硼酸盐矿物质,偶尔还会有雨水滤出。
 Iv冰上起源猜想
Iv冰上起源猜想
上述的所有环境都有一个共同点,就是热量。我们知道,温暖的环境可以帮助推动化学反应,有些科研人员偏要反其道而行之,他们认为生命可能起源于冰块。当水结冰时,溶解在水中的所有物质都会被脱水,并浓缩成盐水。这些盐水被困在冰的狭窄裂缝中,进而形成聚合物链,低温还能帮助生物分子保持结构稳定,让它们得以继续生长。
为了证明这个大胆的猜想,有科研人员将溶液中的成分与金属离子冻在一起,尝试制造RNA链。他们使用环境中常见的离子,帮助促进催化反应。
实验发现,反复的冻融循环有助于酶的生成,酶能催化化学反应产生更多的RNA链。这非常符合我们最初对生命起源的期待。可见,冰似乎是一种有效媒介,它可以促进生物分子转换,使分子变成一个可以自我繁殖的系统。
遗憾的是,上述猜想都面临了同一个问题,它们无法对新陈代谢如何参与生命进程作出解释,这导致了所有猜想都无法回答一个关键问题,即早期生命的三大要素:遗传物质、新陈代谢和膜是怎样共同作用的。
V 火山口湖猜想
 所有人都曾认为,我们在研究生命起源时需要拆分所有元素,再逐一研究。之前大多数研究人员的研究重点,要么在遗传物质中,要么在新陈代谢上。如今,萨瑟兰的团队正在探究一种新的猜想,认为可能生命起源是“一蹴而就”的,并开始寻找可以让所有元素一起发生反应的化学物质。
所有人都曾认为,我们在研究生命起源时需要拆分所有元素,再逐一研究。之前大多数研究人员的研究重点,要么在遗传物质中,要么在新陈代谢上。如今,萨瑟兰的团队正在探究一种新的猜想,认为可能生命起源是“一蹴而就”的,并开始寻找可以让所有元素一起发生反应的化学物质。
萨瑟兰把眼光投向了陨石坑。在40亿年前,陨石撞击地球是很常见的。这些来自外太空的岩石带来了氰化氢,其中包含了有机分子的三大重要成分:碳、氮、氢。如果积聚的氰化氢在水和紫外线光的作用下变热,也许可以形成一整套前驱分子,这些分子可以直接用来合成RNA、蛋白质和脂质。不过这个猜想也有一个问题,就是没有生命把剧毒的氰化物作为碳或者氮的来源。
那么,生命起源的摇篮究竟是湿的、干的还是冷的呢?其实所有的猜想并不是相互排斥的。比如在萨瑟兰的陨石猜想中,至少需要一个干湿循环过程和硫化氢的参与,这两者都可以在陆地起源猜想和海洋起源猜想中寻找帮助。当一颗巨大的陨石撞击地球时,会使地壳裂开。
这时如果在陨石坑中有水池,它可以通过地壳裂缝渗透下去,到地球内部足够深的位置时,又会被再次加热再向上冒泡。年轻的地球大陆上有很多火山口湖泊,它们有一些是由陨石撞击后形成的,这些湖泊的边界还会结冰,就像冬天的美国黄石公园的景致。在边缘的冰和附近陆地的碱性热泉喷口里,我们可以找到RNA中的酶以及新陈代谢作用。
 陨石坑本来就是一个复杂的环境,陨石坑表面的矿物可以充当催化剂,有机物小分子可能会交替溶解在水中并在阳光下干燥。铁镍陨石很容易形成生命起源需要磷酸盐和铁。而且陨石坑还有一个优势:陨石撞击会冲击大气,产生氰化物,形成生命起源的原始有机分子。
陨石坑本来就是一个复杂的环境,陨石坑表面的矿物可以充当催化剂,有机物小分子可能会交替溶解在水中并在阳光下干燥。铁镍陨石很容易形成生命起源需要磷酸盐和铁。而且陨石坑还有一个优势:陨石撞击会冲击大气,产生氰化物,形成生命起源的原始有机分子。
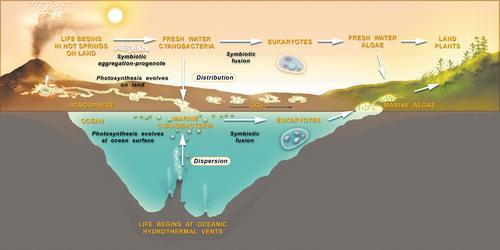
地球早期生命演化之旅
vi 生命到底起源于何处
生命到底起源于何处,恐怕科学家只能依靠理论的推测,随着地球生态系统的复杂化,人类无法模拟出四十多亿年前的那个朴素的地球,只能通过盲人摸象的方式拼凑出生命出现的种种可能,形成一条可行的证据链。
虽然我们喜爱灵光一闪的故事, 喜欢那些能改变世界的重大发现,但更多的时候科学是一种深耕,需要依靠科学家小心翼翼地剔除那些失败的假设,直到真相显露。例如,生命不是起源于海洋,而是经过很多年后,最终被水淹没成了海洋。
目前为止,科学界普遍承认的原初生物留下的遗迹主要有两个,即原初的RNA世界和细胞内的还原环境。
在现代的细胞中,绝大多数化学反应是由蛋白质来催化的,但是蛋白质自身的合成,却仍然要由RNA来催化。组成蛋白质的肽链是在核糖体(ribosome)中合成的,其中的蛋白质亚基只起结构和调节的作用,真正把氨基酸连到肽链上,使肽链延长的,是其中的RNA分子。RNA既能够催化自身的形成,也能够把氨基酸连到小RNA分子上(即后来的转移RNA,tRNA),再把这样带“标记”的氨基酸连接到不断伸长的肽链上。RNA中核苷酸的序列,像DNA中的脱氧核苷酸序列一样,也能够用来储存信息,即为蛋白质分子中的氨基酸序列编码。就是细胞“剪接”RNA以除去内含子(intron)的剪接体(splicesome),也是由能够自我剪接的第II型内含子(RNA)演变而来的。这些事实都说明,最初的生命是RNA的世界,蛋白质是后来才发展出来的。
在原初生命形成时,大气中还没有氧气,而主要由中性气体(如氮气)和还原性气体(如氢气、氨和硫化氢)组成。在此环境中形成的细胞,内部是高度还原的。在这种环境下形成的蛋白质,特别是其中的酶,也只能在还原环境中才能最好地工作。这种情形一旦形成,就难以改变。大气中的氧气出现在大约22-24亿年前,从此大部分生物的环境转变为氧化性的。为了保持细胞内的还原环境,细胞内普遍含有还原性分子如谷胱甘肽(glutathione,浓度大约5 mM),它使得蛋白质分子中的半胱氨酸残基的侧链保持在还原状态,即不形成二硫键(两个巯基-SH 被氧化过程连成-S-S-键)。后来变为叶绿体的原核生物蓝细菌(cyanobacteria)和后来变为线粒体的原核生物a-变形菌(a-proteobacteria)就已经能够合成谷胱甘肽,说明生物很早就发展出对抗环境中氧化状态的能力。在动物体内,在分子中形成二硫键的蛋白或者是分泌到细胞外的,例如抗体分子和胰岛素,或者主要部分位于细胞膜表面的(也即在细胞外),例如胰岛素受体。植物用谷胱甘肽-抗坏血酸循环(glutathione-ascorbate cycle)来消灭细胞内的活性氧物质,维持细胞内的还原状态。现在许多在试管内进行的酶反应,都需要加入还原性的分子如巯基乙醇或二硫苏糖醇(Dithiothreitol,DTT),使反应体系保持在还原状态,使酶能够正常地工作,而不受大气中氧气的影响。
这两大遗迹都有大量事实为根据,而反对的意见基本没有,可以认为是被普遍承认的。问题是,细胞内钾高钠低的状况也是原初生命留下的另一大遗迹吗?
vii 原初生物形成时可能的水环境
钾高钠低的状况存在于所有生物的细胞中,而和地球上几乎所有的液態水中钠和钾的相对浓度相反。无论是河水、湖水还是海水,所含的钠都大大多于钾。例如海水中钠的浓度就是钾的47倍(钠470 mM,钾10 mM),河水中含盐量随河流不同,但大体上钠的浓度是钾的10倍(钠大约0.4 mM,钾大约0.04 mM)。这说明细胞内钾高钠低的状况不是生物在演化过程中“适应”这些水体的结果,而更可能是原初生命形成时环境中的水溶液组成的遗迹。
如果检查一下地壳中钾和钠的含量,发现它们其实差不多:钠为2.8%,而钾为2.6%。之所以河水中钠的含量远高于钾,是因为在岩石风化过程中,钾离子比钠离子更难被释出。例如长石(feldspar,花岗岩的组成成分之一)中的钾就很难溶出。河水入海之后,随着水分的蒸发,盐的浓度也逐渐提高,但是海水的含盐量也不会一直增加下去而变得越来越咸。浅滩处盐结晶出来又被埋藏,形成盐矿;风将大量海水微滴带到陆地,都是减少海水含盐量的方式。另一种除盐的方式是海水通过地壳裂缝与深处高温岩石的相互作用,形成云母(mica)等新的矿物,同时把海水中的金属离子带走。这个过程叫做“反风化”(reverse weathering),是海水中的盐被带走的一个重要方式。据估计,通过这些除盐机制,海水中的盐分在过去的至少15亿年中并没有显著增加。而在反风化过程中,钾比钠更容易被除去,这也使得海水中钠的含量远高于钾。其余的离子,例如钙离子、镁离子、氯离子等,被“反风化”过程除去的情形也不同,因此海水并不是“浓缩的河水”,但是在钠高钾低这一点上,海水和河水是一样的。在有35亿年历史的,位于澳大利亚西部的皮尔巴拉地块(Pilbara Craton)的岩石中,发现了包藏下来的远古海水,其钠离子的浓度高达1 M,大约是现在海水钠浓度的两倍以上。虽然这不一定代表35亿年前海水的平均含盐量,但是也说明远古时期的液体水中就有高浓度的钠离子,生命是不太可能在这样的海水中诞生的。如果说原初生命是在高钾低钠的溶液中产生的,这样的水溶液在哪里呢?
为了解释原初生命产生时水环境钾高钠低的状况,科学家们提出了两种可能性。一种是比35亿年更远古的地壳。这样的地壳在地球上由于板块运动已经难以找到,但是却保留在月球上。其岩石富含钾和磷,叫做KREEP岩石,代表“富含钾(K),稀土元素(Rear Earth Element)和磷(P)”的岩石。由于月球是大约45亿年前一个火星大小的星球和地球相撞形成的,当时的地球应该和月球有相似的地壳,也就是地球早期的地壳很可能也是由KREEP岩石组成的。那时地球的地壳中还没有花岗石(granite),即使在40亿年前,花岗石也很罕见。如果液态水在KREEP那样的岩石上形成,风化过程就应该提供一个富含钾和磷的水环境,十分有利于生命的形成。
另一个可能性是俄裔美国科学家库宁(Eugene V Koonin,1956-)提出的,即热泉蒸汽冷凝所形成的水。钾离子由于比钠离子大得多,比较容易被蒸发的水分子“夹带”,进入蒸汽中。这样蒸汽在冷凝以后,就会形成钾高钠低的水。这个假说也得到了实地观测的证实。例如在意大利的Larderello热泉冷凝水中,钾离子的浓度就是钠离子浓度的32倍。在美国加州的一处热泉,冷凝水中钾离子的浓度竟然是钠离子浓度的75倍!在目前的地球上,这样的热泉为数不多,因为地壳在几十亿年前就大部冷却。但是在地球形成的早期,地壳尚未充分冷却的情况下,应该是很多的。
在现代的热泉中,硫化氢会迅速地被大气中的氧气所氧化,生成硫酸,使得现代热泉冷凝水的酸度极高(pH可以低至0),不适合生物形成。但是在地球早期的大气中,游离氧还不存在,硫化氢不会被氧化为硫酸,生物也就可能在这样的冷凝水中形成。
热泉冷凝水也不是高钾低钠水的唯一来源。由于氯化钠在水中的溶解度几乎和温度无关(例如20摄氏度时为每升35.9克,60摄氏度时为每升37.1克),而氯化钾在水中的溶解度却随着温度升高而升高(例如在20摄氏度时为每升34.2克,和氯化钠差不多,而在60摄氏度时为45.8克,明显超过氯化钠的溶解度),如果一部分海水被隔绝出来,在太阳底下蒸发,在温度较高(例如60摄氏度)时,氯化钠首先饱和,结晶出来。由于氯化钾达到饱和发生在氯化钠之后,所以氯化钠结晶上面的水就会富含氯化钾。这些水如果由于自然的原因流到其它的地方,也会含有较高的钾和较低的钠。这些事实都说明,地球早期出现钾高钠低的水是可能的。
早期形成的细胞膜是不完善的,很可能是由脂肪酸和脂肪醇,而不是磷脂组成的,也就是组成早期细胞膜的脂类分子只有一根碳氢“尾巴”,而不像磷脂分子有两条脂肪酸“尾巴”。这种“单尾巴”的生物膜对于各种离子,甚至像核苷酸那样的巨大离子,都是通透的,证据是由脂肪酸组成的膜可以让外加的核苷酸进入由膜包裹的囊泡内,聚合成为核酸,但是核酸这样的大分子却不能穿过膜,到囊泡的外面。因此在原初细胞中,各种离子的组成,包括钠离子和钾离子的组成,应该是和环境水溶液平衡的。原初细胞中的化学反应也就是在这样富含钾的环境中形成并且不断优化,在这种环境条件下形成的化学反应也就依赖于钾,而不是依赖于钠。如果能够证明细胞最原始蛋白的功能确实需要钾,而不需要钠,就能够为原初生命在钾高钠低的环境中形成的学说提供强有力的证据。
viii 最原始的蛋白质需要钾以执行它们的功能
要证明最原始的蛋白需要钾,首先要找出这些蛋白。为此,Koonin及其同事检查了存在于所有生物的蛋白质,将这样的蛋白质看作是最原始的蛋白。只存在于某些生物,而不存在于其他生物的蛋白则被认为是较后出现的(即生物发生分化后在其中一些生物中出现的)。
这样的蛋白开始比较多,但是随着全部基因组(genome)被测定的生物越来越多,原始蛋白的数量也不断减少(因为只要有新测定的某种生物不含有其中的一些蛋白,这些蛋白就会从名单中剔除),最后稳定在60个左右。这60个左右的蛋白就被认为是生物最古老的蛋白。
检查这些蛋白的功能,发现它们多数与蛋白质的合成,即转译(translation)过程有关,再有就是和DNA有关的酶。这也是可以理解的,因为蛋白合成和DNA信息的读取和修复是生物最基本的生命活动。在这些蛋白中,有若干需要钾离子以实现其功能,但是没有一种蛋白需要钠离子,钠离子的存在甚至会影响其功能。这是原初生命在高钾低钠环境中生成的最强有力的证据。
例如在需要钾离子的蛋白中,有一类是属于所谓的“P-环鸟苷三磷酸酶”(P-loop GTPase),包括转译延长因子EF-Tu(elongation factor thermounstable)和EF-G(elongation factor G)。这些蛋白含有一个专门的天冬氨酸残基用来结合钾离子。这两种蛋白的活性都被钠离子所抑制。
核糖体中真正把氨基酸加到肽链上的酶,肽转移酶(peptidyl transferase)不是蛋白质,而是核糖体中的RNA(ribosomal RNA,rRNA,这是原初生物第一个遗迹,即RNA世界的证据)。如果在试管中除去1价阳离子,肽转移酶就不再有活性。把不同的1价阳离子分别加进反应系统,就会发现使肽转移酶活性恢复的1价阳离子的能力从高到低的顺序是:铵离子(NH4+) > 铷离子(Rb+) > 钾离子(K+) > 铯离子(Cs+),而钠离子(Na+)和锂离子(Li+)没有作用。
因此,合成蛋白质的核糖体需要钾离子才能正常工作,而不需要钠离子。在用体外系统来进行蛋白合成时,通常使用的是兔的网织紅細胞裂解物(rabbit reticulocyte lysate),所需要的阳离子的最后浓度是0.5 mM醋酸镁和79 mM 醋酸钾,而没有氯化钠(根据Promega生物公司的反应系统)。
核糖体合成肽链后,有些还需要伴侣蛋白的帮助才能折叠成为正常的三维结构。其中的伴侣蛋白GroEL也是60个最原始的蛋白之一。它和蛋白GroES一起帮助肽链折叠。其活性受镁离子的帮助,但是绝对依赖钾离子。铵离子和铷离子可以部分取代钾离子的作用,但是锂离子、钠离子和铯离子没有作用。GroEL和GroES都是原核生物的蛋白质,在真核生物中,对应的蛋白质分别是热休克蛋白Hsp60和Hsp10,它们也需要钾离子。这说明从原核生物到真核生物,这些古老的伴侣蛋白和它们的后继物都需要钾离子才能正常工作。
细菌的RecA蛋白、古菌的RadA蛋白、和真核生物的Rad5蛋白都是修复DNA双链断裂的蛋白质,属于最古老的60种蛋白。它们的活性除了需要镁离子外,还需要钾离子,而钠离子没有作用。
CDP-二甘油酯合成酶(CDP-diglyceride synthase)是合成磷脂的重要酶之一,属于60个最古老的蛋白质。它的活性也依赖钾离子,而其活性被钠离子所抑制。
除了为蛋白质的功能所需,钾离子也对细胞膜的形成有利。实验证明,在离子浓度增加到一定程度时,细胞膜会沉淀出来。在没有二价离子(例如镁离子和锰离子)的情况下开始使细胞膜沉淀出来的钠离子浓度(0.4 M)远比钾离子(大于1 M)低。即使在二价离子存在的情况下,使细胞膜凝聚的钠离子浓度仍然比钾离子低,也就是细胞膜在钾离子的环境中更稳定,更容易存在于溶液中。在日常生活中我们也有这样的经验:钾肥皂是液态的,而钠肥皂是固态的。
所有这些事实都表明,生物一些最古老的蛋白质(以及核糖体RNA)的活性需要钾离子。钠离子不仅不能使这些蛋白进入工作状态,在有些情况下还抑制它们的活性。钾离子也有利于细胞膜的生成。这些都是原初生物在高钾低钠的环境中形成最好的证明。这也是为什么地球上的生物在形成并演化几十亿年之后的今天,细胞仍然要保持内部钾高钠低的环境,尽管细胞外的环境几乎全是钠高钾低的。
IX 生命诞生于陆地
现代生物使用纯DNA携带它们的基因,但纯DNA最初可能并不存在。它们可能是RNA核苷酸与DNA核苷酸的混合体。哈佛大学医学院教授索斯塔克证明,这种混合体可以构成“镶嵌”分子,其外貌和行为都很像RNA。这些RNA与DNA混合链甚至可折叠。这表明,最早的生物是否能制造纯RNA或纯DNA都不重要,它们可以使用混合版的RNA,甚至混有TNA或PNA的核苷酸。这不是RNA世界,而是“大杂烩世界”。
这些研究显示,制造最早的细胞似乎并不太困难。细胞的确拥有复杂的机制,但事实证明,它们可以吸收任何东西维持自身生存,虽然这依然不是很好。这种粗细胞似乎不太可能在地球早期生存下来。但当时没有太多竞争,也没有具有威胁性的掠食者,为此从多方面来看,它的生存环境比现在容易得多。但是苏瑟兰德和索斯塔克的理论也存在缺陷,第一种生物肯定拥有某种新陈代谢机制。
从一开始,生命就必须获得能源,否则它无法生存。即使马丁和拉塞尔有关生命起源于深海热液喷口的理论存在错误,但其部分元素肯定是正确的,比如金属对生命起源非常重要。在自然界,许多酶的核心处都有金属原子,这通常是酶的活跃部分,分子的其他部分基本属于支持结构。第一个生命没有这些复杂的酶,为此它们很可能是用“裸金属”作为催化剂。这样看来,热液喷口变得更加重要。如果你看到现代新陈代谢,看起来真的很像铁簇。它与生命诞生于喷口中或附近的理论不谋而合,因为这里富含铁和硫。如果苏瑟兰德或索斯塔克的理论正确,喷口理论就证明是个错误,生命不可能起源于深海。苏瑟兰德说,我们发现的化学物质十分依赖紫外线,而紫外线的唯一来源就是太阳,为此其反应只会发生在有阳光照射的地方,这就排除了深海喷口理论。索斯塔克也认为,深海并非生命温床。但这些问题不能完全驳斥热液喷口理论,或许喷口位于浅水中,那里有阳光,氰化物也可接触到。
阿尔缅(Armen Mulkidjanian)则提出新的假设,生命或许起源于陆地,比如火山口中的池塘中。无论细胞属于哪种生物,它们都含有许多磷酸、钾以及其他金属,但很少有钠。如今,细胞可能通过泵出或泵进机制实现这个目标,但第一个细胞可能无法做到,因为它们不具备这种必要机制。阿尔缅认为,第一个细胞肯定形成于与现代细胞拥有相同化学物质混合物的地方。这立即就排除了海洋,因为细胞中所含磷钾比例远高于海洋,但钠却更少。而活火山附近发现的地热池塘却更为理想,这些池塘中拥有细胞中发现的所有金属物。索斯塔克还认为,地热活跃区的浅湖或地表池塘也很理想,比如黄石公园火山区的那种热液喷口。苏瑟兰德的化学物理论在此也可有很好的解释。温泉中有合适的化学物质,水位波动会导致某些地方干涸,还有来自太阳的大量紫外线。索斯塔克认为,这些池塘同样适合他的原细胞诞生。
X 早期生命的第一个代谢酶推测生存环境
2020年,罗格斯大学的研究人员发现了人体一种蛋白质结构的起源,这种蛋白质结构在人体内负责新陈代谢,这些简单的分子为地球的早期生命提供了能量,并且还可以充当化学信号,研究发表在《美国国家科学院院刊》(Proceedings of the National Academy of Sciences)杂志上,可预测35亿到25亿年前最早的蛋白质是什么样子。
像一个千变万化的谜一样,科学家们追溯了酶(蛋白质)从古老过去到现在的进化过程。解决这个谜需要两块缺失的碎片,没有它们,地球上的生命就不可能存在。通过构建一个由它们在新陈代谢中的作用连接起来的网络,这个团队发现了这些缺失的部分。
该研究的合著者、罗格斯大学罗伯特·伍德·约翰逊医学院(Rutgers Robert Wood Johnson Medical School)生物化学和分子生物学教授、高级生物技术和医学中心(Center for Advanced Biotechnology and Medicine)常驻教员维卡斯·南达(Vikas Nanda)说:“我们对地球生命的起源知之甚少。这项研究可以让我们跨越时间,对最早的代谢蛋白提出假设,我们的预测将在实验室中进行测试,以更好地了解地球生命的起源,并且启发生命在其他地方起源的可能方式。我们正在实验室里建立蛋白质模型,并测试它们是否能引发对早期生命新陈代谢过程至关重要的反应。”
在美国国家航空航天局的资助下,由罗格斯大学领导的一个名为ENIGMA(Evolution of Nanomachines in Geospheres and Microbial Ancestors)的科学家团队正在通过美国国家航空航天局天体生物学计划的成员身份进行这项研究。ENIGMA项目试图揭示最简单的蛋白质在生命早期阶段的催化作用。
资深作者、ENIGMA首席研究员和罗格斯大学新伯朗士威校区杰出教授、环境生物物理学、分子生态学实验室负责人保罗·G·法尔(Paul G. Falkowski)说:“我们认为生命是从非常小的基石进化而来的,并且像乐高积木套装那样,发展出细胞和像我们一样更复杂的生物体。我们认为我们已经找到了生命的‘积木’——正是这些‘乐高积木’最终导致了细胞、动物和植物的进化。”
罗格斯大学研究小组着重研究了两种蛋白质折叠,它们可能是早期新陈代谢的第一个结构。这两种蛋白质折叠分别是铁氧还蛋白折叠(ferredoxin fold)和“罗斯曼”折叠(Rossmann fold),前者与铁硫化合物结合,后者与核苷酸(DNA和RNA的基石)结合。这就是生命进化过程谜题中缺失的两个部分。
从最早产生的蛋白折叠中我们可以看到,最早的生命代谢是在富含铁和硫的环境中产生的,铁氧还蛋白是含有铁原子和无机硫化物,具有电子传递体作用的小分子蛋白质,它存在于现代蛋白质当中,并且穿梭在细胞周围的电子里以促进新陈代谢。电子会在固体、液体和气体中流动,为生命系统提供动力。
rId13
所谓折叠模式,就是它们包纳反应物的结构,具有特异性,一种折叠模式可能只对应着少量的化合物。“罗斯曼”折叠是与核苷酸(DNA和RNA的基石)结的结构,证明最早产生的蛋白质只干两件事情,催化铁-硫产生能量,硫化合价降低被还原,铁化合价升高被氧化,即使被氧化成亚铁离子,也能产生两个自由电子,助推生命反应。
而富含铁硫矿的地方,只能在火山口、陨石坑或者铁硫矿床寻找。
第三章 生命起源比我们想象的要早
目前,地球仍然是宇宙中已知唯一拥有生命的地方。地球的生物圈延伸到地表以下至少19公里,并延伸到大气中至少包括土壤、热液喷口和岩石 ,甚至在南极冰层以下800米处。包括海洋最深处,直到海底以下几公里的岩石。研究显示,可以在任何地方发现微生物,它们非常适应条件,无论在哪里生存。在某些测试条件下,已经观察到生命形式可以在外层空间的真空中生存。
在地球上曾经生活过的所有生命形式中,超过50亿即估计约超过99%的品种已灭绝。地球目前物种数量的估计数在1000万至1400万之间,其中大约120万被记录,而超过86%的数据仍未描述。 研究表明,地球上的生命可能来自太空尘埃或陨石所携带的生物物质。地球上发现的最古老的陨石碎片大约有45.4亿年的历史。研究表明,地壳在行星形成后大约1亿年就凝固了,并且行星迅速获得了海洋和大气层 ,这可能已经能够维持生命。
根据化石记录,现在普遍认为地球形成于45.4亿年前,在1.3亿年后,也就是44.1亿年前,地球上首次出现了海洋。又过了1.3亿年,在42.8亿年前,地球上首次出现的生命。但是我们认为生命可能出现在海洋之前。无论是海洋出现的速度还是生命出现的速度,都比我们想象中要快得多。从原始地球形成到首次生命的出现,整个过程不超过3亿年。对于这个充斥生命的星球,我们可能早已习以为常了。可是你是否曾经有那么一瞬间沉下心来认真思考过,地球上的第一个生命到底是怎么来的呢?
二十世纪初,苏联生物化学家 Alexander Oparin 和英国遗传学家 J. B. S. Haldane 分别提出“原始汤”假说,认为早期地球的海水里的碳基化学物质互相反应,形成蛋白质、核酸等“生命的基础”。
1953 年,Stanley Miller 在模拟远古地球大气的气体中放电,发现产生了少量的氨基酸等有机物。可是,蛋白质和核酸并不能在水中长期稳定地存在。细胞质是一团胶体,性质与海水截然不同。这意味着细胞生物自组织起来的环境需要水且水不能太多——陆地比海洋更适合。
a Sutherland 等成功地从磷酸盐和四种简单的碳基化学物质开始、在模拟远古地球的环境里造出两种核酸。反应过程中,化学物质高度浓缩地溶解在水中,且一些步骤需要紫外线辐射,显然无法发生在海洋里——暴露在太阳紫外线辐射下的水坑等小规模水体才能满足要求。
b那之后,他们略微改变反应条件,产生了蛋白质和脂质的前体。
2019 年,Carell 团队搞出了能在远古地球的环境条件及简单的无机底物作用下同时产生四种RNA核苷酸的过程。该过程不需要复杂的分离和纯化,即可产生关键的生命组分:
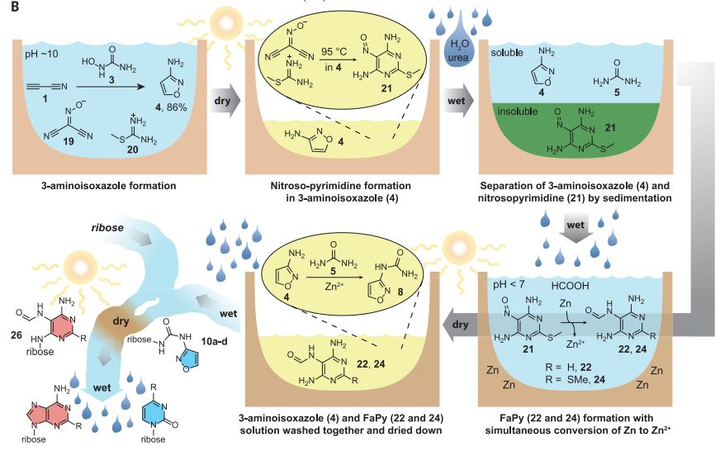
相比之下,化学家尚未在模拟海水的条件下合成这些分子。
Moran Frenkel-Pinter 等证明,氨基酸在没有水的条件下可以自组织成链,地球生物使用的蛋白氨基酸比非蛋白氨基酸更容易发生这种反应。这可以帮助解答为何地球生物使用这些氨基酸来组成蛋白质。
2018 年,分子钟显示地球生物的起源可能早于地球上海洋的形成。
目前的理论下,地球似乎是在 45.4 亿年前形成的,冥古宙由此开始。
在 45.2 亿年前,地球似乎与体积约等于火星的天体“忒伊亚”发生碰撞,飞溅出的物质有一部分形成了月球,地球从炽热的岩浆球状态逐渐冷却固化(计算表明需时1亿年)。
44.1 亿年前,地球上可能出现了原始海洋。这一时期的地质活动估计相当剧烈,火山喷发遍布地面、熔岩四处流动。
在 41 亿年前到 38 亿年前,地球可能受到了大量小行星与彗星的撞击。根据同时期的月球撞击坑推算[2],地球当时形成了 22000 个或更多的直径大于 20 千米的撞击坑、约 40 个直径约 1000 千米的撞击盆地、几个直径约 5000 千米的撞击盆地,地形平均每 100 年就受到显著破坏。
冥古宙在 38 亿年前结束,内太阳系不再有大规模撞击事件。
也有研究认为上述阶段的撞击规模要小一个甚至几个数量级。
21 世纪初,学者一般估计现存生物的最后共同祖先(LUCA)生活在距今 35 亿年前~38 亿年前,而我们发现的直接的古生物化石证据已经老到了 34.8 亿年前[3]。
2017 年,科学家在加拿大魁北克的岩石中发现了 37.7 亿年前~42.8 亿年前的筒状微小纤维构造,可能是远古海底热泉喷口处生物的活动痕迹[4]。
2018 年,有研究根据分子钟将最后共同祖先生活的年代设置到了 45 亿年前[5]——这意味着 LUCA 可能在地球形成后 4 千万年时地狱般的环境里就自然形成了,比海洋的形成早九千万年。
地球生物的太空起源说仍然是学术界经常谈论的话题,人们实际上发现一些陨石可以带来大量的氨基酸等有机物,并在撞击时产生一个灼热的坑,该坑的任意边缘与水域相接就可以在坑里灌上水。
研究人员从当地时间 1969 年 9 月 28 日坠落在澳大利亚的默奇森陨石中找到了 70 种以上的氨基酸,既有常见的甘氨酸、丙氨酸、谷氨酸之类,也有不寻常的异缬胺酸、柳氨酸之类,有些取代基有 9 个碳原子,还有二羧基、二氨基等不同的官能团。2020 年 1 月发表的一项研究估计该陨石夹带的碳化硅颗粒有 70 亿年的历史。


来源:阿贡国家实验室
科学家对在南极发现的陨石进行的更多研究显示,富含碳的陨石坠落时的高温可能驱动化学反应产生大量的氨基酸。
2022 年,实验显示,水溶液中的酮酸、氰化物、氨、二氧化碳可进行化学反应,产生乳清酸盐和氨基酸。乳清酸是核苷酸的前体。这种反应可以在原始地球的条件下进行,而且需要的水量很少。
文章发表在Nature Chemistry 28 July 2022, .DOI: 10.1038/s41557-022-00999-w
据最新一期《科学》(2024.2.26)杂志报道,英国伦敦大学学院研究团队在实验室中成功合成了一种对生物体至关重要的化合物——泛酰巯基乙胺。这一发现表明,该化合物可能在地球早期就已存在,并在生命诞生之初发挥了关键作用。
地球生物也不必都是在同一个时代起源的,现代地球的水域、地下仍然可能在进行有机大分子的自我制造,只是这个过程本就极度缓慢、难以和现代生物制造的有机大分子区分、很容易和现代生物的身体结合到一起去(例如被细菌摄入体内)。
45.7亿年前,太阳诞生,但构成我们的星际物质,或许百亿年前就已经存在。
45.4亿年前,地球诞生。
44.5亿年前——地球与忒伊亚大碰撞,月球形成。
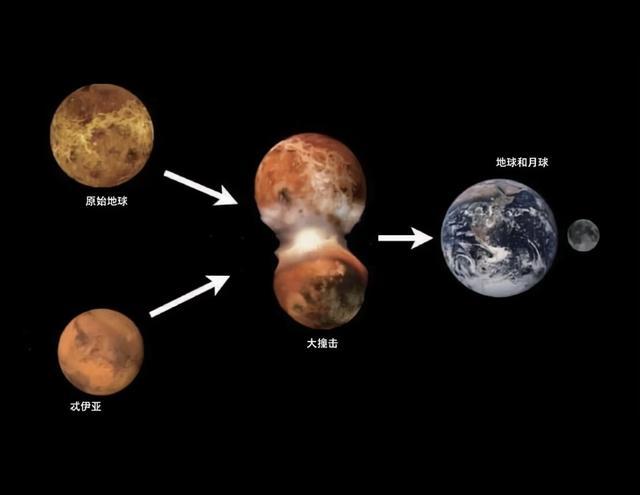
44.9亿年前——地月潮汐锁定。
44亿年前——第一滴液态水出现。
42.8亿年前——或许已有了最早的生命。
41亿年前——地球遭遇超级流星大轰炸,一轰炸就是3亿年,或许带来了生命物质。
讨论最原始生命起源的基本问题时,生物学和化学之间的界限就不再存在。非生命向生命的过渡是连续的。就像一个连续的色谱,可以在其中指出某些地方是蓝色的,某些地方是绿色的,但也有一些地方是蓝绿色的,难以完全地确定颜色。
当科学家试图定义生命的起源的是什么?为什么?怎样的?等基本问题时,也是如此,没有完美的定义,可以有众说纷纭的标准。但是,大多数科学研究都采用约定的概念:生命是复制和进化的结果。生物学家认为,地球上所有活的生物必须共享一个最后的祖先 ,因为两个或多个独立的世系不可能独立地开发出所有活生物共有的许多复杂的生化机制,这几乎是不可能的。
研究表明,地球上的生命是基于碳和水的结果。碳为复杂的化学物质提供了稳定的框架,可以轻松地从环境中提取,尤其是从二氧化碳中提取。没有其他化学元素的性质与碳相似可以称为类似物。硅是元素周期表中碳正下方的元素,不会形成很多复杂的稳定分子,并且由于其大多数化合物是水不溶性的,因此生物体更难以提取。硼和磷元素的化学性质较为复杂,但相对于碳还具有其他局限性。
水是一种极好的溶剂,并具有其他两个有用的特性:浮冰使冬天的水生生物能够在其下生存;其分子具有负电和正电端,这使其能够形成比其他溶剂更广泛的化合物。其他良好的溶剂(例如氨)仅在如此低温下呈液态,以至于化学反应可能太慢而无法维持生命,并且缺乏水的其他优势。
在有机的、水基的地球生命中,我们有两种途径来回答“最原始的生命形式”这一问题:一种是看近代最原始的生活体系,另一个是假设原始生命形式在地球早期会是什么样子。
对于第一种途径,可以将细菌视为原始的生命形式。但是,它们存活的环境是另一个生命系统。有细菌依赖其他细菌存活。但最原始的生命形式取决于另一个存活的生命系统。另一个可能的答案是指向最近宣布的能够在营养丰富的环境中生活的合成细胞,该细胞具有迄今为止任何已知的独立复制生物中最小的基因组。
对于第二种途径,生命起源中最常见的理论表明,在开初的化学反应中的某个时刻,出现了复制酶。复制酶是可以自我复制的分子,也可以复制其他相似的分子。核糖核酸(英语:Ribonucleic acid,简称RNA)是最适合在早期地球上发挥复制作用的化学物质。 RNA是一种非常类似于DNA的聚合物,可以存储遗传信息,但也可以折叠成酶促结构,并有助于催化化学反应。
对于非生命化学物质如何产生生命的研究,集中在三个可能的起点上: 自我复制 ,一种有机体产生与自身非常相似的后代的能力; 新陈代谢,其养活和自我修复的能力; 和外部细胞膜 ,允许食物进入和浪费产品离开,但排除不需要的物质。关于生物发生的研究还有很长的路要走,因为理论和经验方法才刚刚开始相互联系。
即使已发现具有令人印象深刻的功能的催化RNA,但距离找到足够好的可自我复制并具有足够高性能的复制酶,还有很长的路要走。要拥有高效的复制,需要一个复杂的分子,而要良好地复制复杂的分子,需要一个高效的复制。
显然复制其他更简单的分子比自我复制更容易。但这存在一个问题:寄生虫,那些被复制但从不自我复制。这些寄生分子也可能由于复制错误(称为突变)而出现。一旦出现在大量复制者中,便会迅速吞噬复制者使用的资源,并导致其他生物灭绝。
即使是三个现代生活领域中最简单的成员,也都使用DNA来记录其“ 配方 ”,并使用复杂的RNA和蛋白质分子阵列来“读取”这些说明并将其用于生长,维持和自我复制。 一些RNA分子可以催化自身复制和蛋白质构建的发现导致了完全基于RNA的早期生命形式的假说。 这些核酶可能形成了一个RNA世界 ,其中有个体但没有物种,因为突变和水平基因转移将意味着每一代后代的基因组很可能与其父母最初的基因组不同。 RNA稍后将被DNA取代,DNA更稳定,因此可以构建更长的基因组,从而扩大了单个生物体具有的能力范围。 核酶仍然是核糖体 (现代细胞的“蛋白质工厂”)的主要成分。
尽管在实验室中人为地制造了短的自我复制RNA分子,但是人们怀疑是否可以进行天然的非生物合成RNA。 最早的“核酶”可能是由较简单的核酸形成,后来被RNA取代。
在存在寄生虫的情况下,如何维护复制品?答案是复制品可以合作。如果复制品在膜(可能由脂质制成)的帮助下粘在一起并形成基团,那么它们比容易与寄生虫混合更好。膜内部的合作分子将由于自身或彼此的产生而无寄生虫地生长,并最终导致膜分裂。
所以,最原始的生命形式(在地球上可以想象到)包括组织起来相互协作的元素,很可能是在分裂的膜内。最可能合作的形式涉及复制其他分子。最原始和最早的生命形式是一种膜,可能包含完整的分裂循环,而其中包含协作元素。
最早形成生命的方式最早出现在地球上至少是37.7亿年前,可能最早是42.8亿年,甚至是45亿年。在44.1亿年前海洋形成之后不久,在45.4亿年前地球形成之后不久。据研究报告指出,34.65亿年前的澳大利亚安培克思Apex石曾经含有微生物 ,这是地球上生命的最早直接证据。 在西澳大利亚34.8亿年前的砂岩中发现了微生物垫化石。在格陵兰西南部37亿年前的沉积沉积岩中发现了生物石墨以及可能的叠层石的证据,在西澳大利亚州的41亿年前的岩石中发现了“生命遗骸 ”。
但是随着对生命起源研究的深入,许多科学家发现生命起源于海洋的假说存在一个根本问题:构成生命基石的有机大分子可以在水中分解,蛋白质和核酸的链接处很容易受到大量水分子攻击而断裂。
水悖论
1986年,已故的生物化学家罗伯特夏皮罗在其著作《起源》中写道:在碳化学世界中,水是大分子最大的敌人,因为分子可以被水分解,这就是水悖论。
在生物体内,细胞通过限制水在细胞内的自由流动来解决分子被水分解。因此,流行的细胞质图像通常是错误的。生物课本中描述的细胞基质只是一个容纳所有东西的袋子,所有东西都可以游动,这并不正确,细胞基质是凝胶体而不是单纯的水,生物必须控制水的通量。
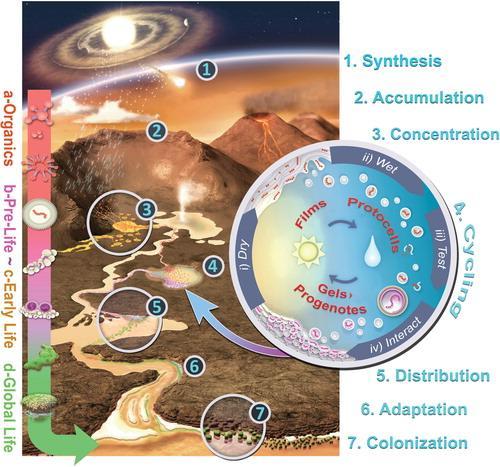
因此,广阔的海洋环境可能并不利于生命形成,它无法使化学物质浓缩。生命的关键分子及其核心过程只能在相对较浅的水域中形成,水环境必须高度浓缩,甚至有时会完全变干,在干湿交替的陆地环境中,生命才可以形成。
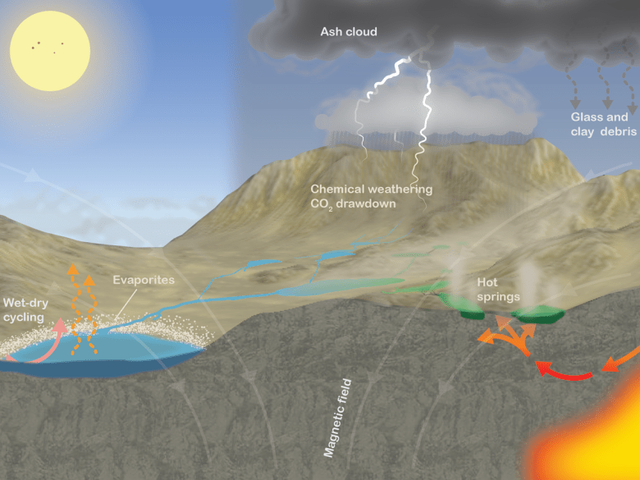
2009年,科学家找到生命起源于干湿交替环境的证据。科学家萨瑟兰成功地制造出构成RNA四个核苷酸的其中两个。他将磷酸盐和四种简单的有机物溶解在水中,但高度浓缩,通过加热这些物质,使其经受紫外线辐射并间歇性干燥,最终结果产生RNA、蛋白质和细胞的其他核心成分,而在此之前这被认为是不可能的,同时在之前的生命起源于原始海洋的实验中从未合成过如此广泛的生物分子。

干湿交替的环境
所以,萨瑟兰认为生命不可能在海洋深处起源,只能在阳光充足的池塘或溪流中,组成生命的简单有机物才能集中。
2019年,美国生物化学家皮特对这种观点进行扩展。他们证实如果氨基酸变干,氨基酸会自发连接形成蛋白样链。与地球古老的其他氨基酸相比,现在蛋白质中存在的20多种氨基酸更有可能发生这类反应。
囊泡封装核糖
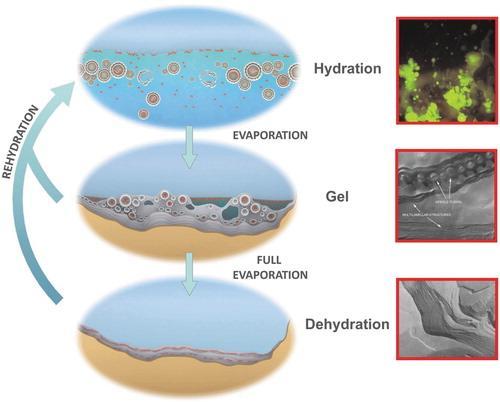
同时,间歇性干燥也可以驱动大分子构件组装得更复杂。1982年,加利福尼亚大学科学家研究了大分子脂质如何自发形成包围细胞的细胞膜。他们首先制造囊泡:即球形斑点,其亲水基核心被两个脂质层围绕。然后研究人员将囊泡干燥,脂质重新组织形成多层结构,就像一叠煎饼。先前漂浮在水中的DNA链被困在两层之间。当研究人员再次加水时,囊泡进行重整,DNA进入囊中。
2008年,科学家将核苷酸、脂质与水混合,然后进行干湿循环。当脂质形成层时,核苷酸连接成RNA状链。
除此之外,这些研究指出生命起源的另一个关键因素:光。
合成生物学家杰克·索斯塔克(Jack Szostak)利用一些简单细胞——即包含少量化学物质,但可以生长,竞争和自我复制的细胞进行一些实验。如果将这些原始细胞暴露在与陆地相似的条件下,它们表现出更逼真的行为,原始细胞可以利用光能以简单复制形式进行分裂。
其次,紫外线辐射可以推动铁硫簇的合成,这对许多蛋白质合成至关重要,铁硫簇的合成可以驱动储能分子ATP的合成,有助于为活细胞提供动力。但是如果将铁硫分子暴露在水中,它们就会破裂。
因此,许多科学家相信生命始于光线充足,水量有限的陆地表面。但是,水在生命起源的过程中扮演的角色仍存在争议。燥的环境为蛋白质和RNA等分子的形成提供了机会。但是,简单地制造RNA和其他的大分子并不是生命,物质必须形成一个自我维持的动态系统。
生命进入海洋
有科学家认为,水破坏可能会促进生命形成,就像捕食动物比被捕食动物进化得更快,第一个生物分子可能已经进化出应对水的化学攻击的能力,甚至能利用水的破坏性。
在新西兰罗托鲁瓦附近温泉中进行的一项研究表明,来自温泉的样品经历了干燥再润湿的循环之后,产生类似RNA的分子。干燥可以导致氨基酸自发连接。
但是,研究小组也发现它们的原蛋白可以与RNA相互作用,两者在水中都变得更加稳定。实际上,水扮演自然选择的角色,只有那些可以在水中生存的分子可以存在,其他分子则会被破坏。在每个润湿周期中,链接较弱的分子或无法通过结合而自我保护的分子被水破坏。而较稳定的更复杂的分子则不断积累。
所以,生命起源环境中的水不应该那么多,以至于生物分子被破坏得太快;但也不至于没有那么少,以至于生物分子没有任何变化。
据推测,44亿年前或者更早——第一滴液态水出现。在此之前,都是火山喷发的水蒸汽(原始大气最多的是氮气、二氧化碳、水蒸气),生命有机物质、生命的基石和养料在此阶段可能已经完全具备,因为要大量产生原始细胞,就需要大量原料。
第四章 亿万斯年,一场大雨
地球在刚形成的时候看起来是什么样子?这是个发生在人类将自然奇观镌刻石头之前,在树木将四季刻入年轮之前,是山脉经由板块运动突出地表之前,也是山川暴露于古老地层以前的问题。
地球诞生于原始太阳系的尘埃云盘之中,历经星尘、烈火、汪洋、沙漠、寒冰的无数磨难淬炼,终于进化成如今蓬勃灿烂的模样。46亿年里,地球不断变化着,呈现出迥然不同的多副面孔:满是岩浆流淌的熔化地球、布满陨石坑的蜂窝地球、硫化气体熏蒸的恶臭地球、铁锈遍布的玄黄地球、玄武岩覆盖的黑色地球、汪洋无际的蓝色地球、冰盖包裹的白色地球、沙漠绵延的棕色地球、植物疯长的绿色地球……直达今天海陆相间、地貌多样、生机勃勃的生命地球。
为了书写方便,我们简单把地球分为以下几个时代:混沌时代、熔浆时代、滚石时代、暴雨时代、浅水时代、玄黄时代、洪荒时代、深水时代分别论述。
I 混沌时代
最初,没有地球,没有太阳。我们的宇宙也不是从真空中突然出现的,因为在大爆炸之前,连真空和时间也没有,甚至连可以容纳事物的“空”都不存在。一片虚无,一无所有,然后,就有了。宇宙大爆炸——这一创世的奇点,是从无到有的转折,超越了现代科技的眼界,超越了数学的逻辑,但却具有决定性。全部的空间、能量与物质都从不可知的虚无中诞生。
在大爆炸之后的一秒钟的一瞬,最初的亚原子粒子出现了——电子和夸克,这是我们世界中全部固体、液体和气体的本质。此后不久,仍然是在第一秒钟的一瞬,夸克结成对子或三联体,形成质子和中子,落户在每个原子的核中。此时宇宙烫的荒谬,这种极热持续了约50万年,直到持续的膨胀最终把宇宙的温度降到了几千度——这就足够“凉快”了,电子就能被吸附在原子核周围,形成第一批原子。第一批原子绝大多数是氢——超过全部原子的90%,还有百分之几的氦,以及一小撮锂混进来。这些元素的混合物构成了第一批恒星,最早分子HeH3+已经过科学家证明。
科学家认为,在大爆炸的10万年后,中性氦原子(He)与质子(实际上是带正电的氢离子,H⁺)会开始反应,形成宇宙中的第一批分子:HeH⁺(氦合氢离子)。这是宇宙演化的第一步。原子有了、质子有了、分子有了,连恒星也有了,万事俱备只欠东风,在非常高的稳度、巨大的压力之下,开启了核聚变的过程,释放大量的能量。恒星因此膨胀了很多倍,爆炸成最初的超星新。
随着第一批恒星的爆炸,宇宙开始变得越发有趣。老恒星炸飞的碎片,成为新生的恒星群的种子,方式是形成新的星云。每一团巨大的星云,由气体和尘埃构成,那其实是此前许多恒星的残骸。每一团新星云所含的铁都比上一次更多,所含的氢都比以前的恒星群更少。在130.7亿年里,这种循环持续不断。随着老恒星产生新恒星,无数亿颗恒星在无数亿个星系中涌现。包括我们的银河系、太阳系。
46亿年前一片巨大氢分子星云的引力坍缩,坍缩的质量大多集中在中心,形成了太阳。其余部分一边旋转一边摊平,形成了一个原行星盘,继而形成了行星、卫星、流星体和其他太阳系小天体。地球最开始由气体、冰粒、尘埃形成的直径为一至十千米的块状物,这些物质经过1000至2000万年的吸积生长,最终形成原始地球。
对于宇宙来说,没有什么物质是亘古不变的。我们居住的这个天体悬浮在宇宙中,围绕着太阳一圈圈的转动。根据化石记载和上一章的推断,地球约诞生于45.4亿年前。在这从古至今的几十亿年的时间中,地球也转了几十亿圈,地球纪元也经历了很多的历史,难以计数的物种是没有留下痕迹或者人类迄今并未发现痕迹的。不过,无法精确测定到地球元年。在地球从无到有,从气体尘埃到形成的那一刻,可称为混沌时代。
II 熔浆时代
在45亿年前的银河系中,大量的尘埃和小行星围绕着早期的太阳旋转,这些转动的物质既有微小的灰尘,也有直径几百公里的小行星。不久,大大小小的物质开始相互碰撞,没有碰碎的空间物质结合在一起,形成了一个岩石体,这就是地球的雏形。
随着越来越多碰撞物的聚集,地球逐渐长大了,其引力场也逐渐变得越来越强,使周围旋转的星际物质越来越快的拉像地球,以更强的力量冲击地球的表面,形成巨大的陨石坑,释放出大量的热。
在强大热量作用下,地球的外层开始融化,形成了一个沸腾的熔岩浅海,还有大量的热被地球内部吸收,埋藏在成吨的不断生长的岩石下面,这样的过程持续了几百万年,直到地球长成现在的样子。
然而这些描述全都是根据推论而来,地球最初数百万年时期的证据并没有留下来。我们的地球是个终极循环器,板块运动不停地把老岩块翻新,而熔岩流变硬就形成新的地景。我们不能以现在的地表样式来刻画地球刚形成时的样子,45亿年前的地球刚形成时,地表很可能和现在完全不同。科学家依希腊神话的冥王之名,将此时期命名为冥古代(Hadean),这名称极为贴切,因为这时的地表不断受到陨石袭击,地面充斥着汨汨流出熔岩的火山,有着炽热炼狱般的景象,如果这个时期你穿越回去,对于生命来说你看到的绝对是一个地狱一般的场景。
到44亿年前,撞击的减少使岩浆海的活动减弱,地球的表面开始冷却,慢慢地,冷凝的岩浆形成一层薄而黑的地壳覆盖着地球,虽然行星撞击和火山喷发不时地把地壳撕开,把炙热的岩浆喷向天空,但是随着撞击的不断减少,冷却的不断进行,地球表面形成了越来越厚的地壳,地球从熔浆进入到陨石乱飞的时代。
III 陨石时代
地球从诞生到40亿年前,是著名的重轰炸期。顾名思义,地球遭到了高频的全球范围的轰炸,炸弹就是大量小行星、陨石与彗星的轰击。
因为地球的地貌已经被后来的地壳运动和风和河流改变,所以一般会参照月球进行研究,因为月球几十亿年前的地质条件都保存完好。根据重轰炸期月球撞击坑推算(月球面对地球的那一面的大部份大型盆地如危海、静海、澄海、丰富海和风暴洋也都是于此一时期撞击形成的),地球当时的场景可以想象一下:
每隔十几年,就会有一次毁天灭地的撞击,撞击坑直径约5000公里,相当于整个中国大小;每隔2年,都会有一次瞬间毁灭全球的撞击,撞击坑直径约1000公里,相当于法国大小;每天,都会有猛烈撞击的发生,撞击坑直径大于20公里,每个都足以毁灭现代人类文明,当时地球完全超出于任何人的想象,这就是重轰炸期。
冥古宙在40亿年前结束后,内太阳系不再有大规模撞击事件。目前已知的地球最古老的岩石是于格陵兰发现的依苏阿绿岩带,由约38亿年历史的沉积层和火山岩脉所组成。位于北美克拉通盖层的艾加斯塔片麻岩及西澳那瑞尔片麻岩层的形成也基本在38亿年前。
在整个陨石时代,地球从一个炽热的岩浆球,地质活动剧烈、火山喷发遍布、熔岩四处流动的时代,对生命来说每天都是灾难。但是,生命就是在这样恶劣的环境中诞生了。让我们这些40亿年后的地球人类复原那时候的地表情况是几无可能的,然而科学家的确从锆石(zircon)这种极度坚固的矿物中找到了一些线索。
锆石结晶几乎坚不可摧,现存的锆石有些已经将近44亿岁了。它们就像是微小的时光胶囊,里头装着非常早期的化学足迹。这基本上是我们窥看地球成形阶段的唯一机会。罗彻斯特大学(University of Rochester)的达斯汀.崔尔(Dustin Trail)在《美国国家科学院刊》(PNAS)上的一份新研究里,正缓慢地厘清孕育最初生命型态的环境状况。
IV、暴雨时代
从冥古宙结束到太古宙开始。地球表面的温度降低,开始冷却凝固。最初的大气可能由水汽、二氧化碳、氮组成,火山爆发所释放的气体形成了次生大气。水汽的蒸发加速了地表的冷却,形成了极为坚硬的以花岗岩结晶为主的岩石,地球岩石自此开始稳定存在并可以保留到现在。
地表的冷却使大气中的水蒸气凝集,水滴以降雨的形式落到地面上,暴雨连续下了成千上万年,雨水流过地球表面,灌满了盆地、陨石坑和自然形成的凹地,顺着溪流流动,河水夹带着各种矿物质和生命分子,百川汇流形成了海洋。这时候海水是酸性的,而且非常热,水温大概有100℃。
锆石结晶由于非常坚固,所以常常能从这个循环过程的极端温度与压力中存活下来,保留了它们初形成时所处环境的线索。研究人员早先利用锆石的氧同位素分析,发现在43亿年前液态水就已覆盖了部分地表,这显示了地球表面在形成后的数亿年就已经冷却了。
达斯汀.崔尔团队找上加州大学洛杉矶分校的高解析度离子微探针(high-resolution ion microprobe)来帮忙,它会对着微小的样本射出一束带电的原子,然后测量弹出来的那些离子。
他们从西澳洲的杰克山岗(Jack Hills)地区收集了超过40亿年前形成的锆石进行检验,每个大约100微米宽,接近人发的粗细。他们把这些古老矿石的化学性质,拿来和更年轻、起源更确定的锆石(可把这些年轻锆石视为地质史的桥梁)互相比较,有助于理解不同的同位素比率,在被检验的古老锆石中,超过半数显示出岩石与水很早就在许多不同环境中互相作用。
有些锆石含有岩石被水蚀化成黏土的化学特征,其他锆石所带的特征则是矿物溶解后再结晶所形成的岩石,例如湖里或海里的角岩(chert)或带状铁矿床( banded iron formations)。还有一些则具有所谓的蛇纹岩化作用(serpentinization,因其带有蛇皮般的纹理和颜色而得名)的特征。在这个过程中,水和富含铁、镁的岩石发生作用而被并入矿物的结构里。最重要的是,这些过程全都创造了能促进早期生化反应的有利基石,成了照亮早期生命的微光。
我们周遭万物和体内世界一度源自星尘,源自那些形成每个分子、矿物乃至今日复杂生物的早期过程。从手机到食物,从古菌到真核细胞,以及你胸中跳动的心脏皆是如此,而科学家才刚刚开始厘清地球的起源。
V、浅水时代
即使现在,地球的表面也不是平整的,而是坑坑洼洼的。原始地球的地表形态有过之而无不及。地球逐渐变冷,地面温度终于降到水的沸点以下,于是倾盆大雨从天而降,如同千军万马,奔向低凹之处。那时的地球,时而在这里,时而又在那里,降着倾盆大雨。如注的雨水,一遍又一遍地冲刷着一向干燥的地面,它们汇成巨流流向低洼的地方,日复一日,月复一月,年复一年。经过长期的降雨,便形成了江河湖泊和汪洋大海。
科学家们把最早形成的大海称作 原始海洋 。此时的大洋水不仅严重缺氧,而且含有大量的火山喷发酸性物质,如HCL、HF、CO2等,具有较强的溶解能力。根据科学家对化石的研究,大约在40亿年前就形成了原始海洋。
原始海洋的规模远没有现代海洋这么大。据估算,其水量大约只有现代海洋的10%。后来,由于贮藏在地球内部的结构水的加入,才逐渐形成了蔚为壮观的现代海洋。原始海洋中的水不像现代海水又苦又咸。现代海洋海水中的无机盐,主要是通过自然界周而复始的水循环,由陆地带入海洋而逐年增加的。可是,原始海洋中的有机大分子要比现在海洋中的丰富得多。
原始大气化学演化过程中所形成的氨基酸、核苷酸、核糖、脱氧核糖和嘌呤等有机分子都随着雨水冲进了原始海洋,并迅速地下沉到原始海洋的中层,从而避免了因原始大气缺乏臭氧层而造成的紫外线伤害。又经过了很长时间,原始海洋中的有机分子越来越丰富(据推测,在原始海洋中有机分子含量达到了1%)是由于当时大气中无游离氧,因此高空中也没有臭氧层阻挡,不能吸收太阳辐射的紫外线,所以紫外线能直射到地球表面,成为合成有机物的能源。此外,天空放电、火山爆发所放出的能量、宇宙间的宇宙射线,以及陨星穿过大气层时所引起的冲击波等,也都有助于有机物的合成。但其中天空放电可能是最重要的,因为这种能源所提供的能量较多,又在靠近海洋表面的地方释放,在那里它作用于还原性大气,所合成的有机物质,很容易被雨水冲淋到原始海洋之中,使原始海洋富含有机物质,这就为生命的诞生创造了必要的条件。
而我们认为,原始生命材料并不是在原始海洋中产生的,或者说是原始海洋组装了生命。地球上已经原本有很多产生生命的质素,例如RNA、DNA和大量辅酶与矿物质,他们随着雨水一起流向了海洋,而且恰巧淹没了最有可能产生生命的地方,也就是原始的陨石坑、硫铁矿床、硝化池、火山口、喷泉、冰面。沙漠等等所有可能产生生命的地方,海洋并不是产生生命的第一现场,而是生命汇集的地方,正好,因为海水漫灌,淹没了,使原本的产地成为了土著,所以真正值得研究的生命第一现场是陨石坑、硫铁矿床、硝化池、火山口、喷泉、冰面、沙漠等等,当然大海也值得研究——第二现场。
vi、玄黄时代
生命如何起源,首先还得看你如何定义生命。在这里,我们要提出我们的假设,生命起源于那些陨石坑、硫铁矿床、硝化池、火山口、喷泉、冰面、沙漠等等著名的土著现场,它们已经是我们耳熟能详的名场面。
首先,地球上确实拥有支持碳基生命产生的所有原料物质。这些原料主要包括:
有机分子:由碳原子构成的有机分子是生命的基础,它们可以形成从简单的碳氢化合物到复杂的DNA分子等各种生物大分子。
主要元素:构成碳基生命的最主要元素包括碳、氢、氧、氮、磷、硫、镁、锰、钾、钠......这些元素在生物体内发挥着不同的作用,如碳是构成生物大分子的骨架,而氮和磷则是构成核酸和蛋白质的关键元素。
碳的特殊属性:碳原子具有独特的化学性质,它能够与其他元素(包括自身)形成稳定的共价键,创造出多样化的化合物。这种特性使得碳成为构建复杂生命系统的理想的原子。
丰富的碳来源:地球上的碳可能来源于太阳系原行星盘形成并变暖后,由星际介质所积累起来的碳,这些碳在地球形成过程中被纳入地壳,成为生命诞生的物质基础。
所以说,地球上的环境和化学元素为碳基生命的产生提供了必要的条件。从简单的有机分子到复杂的生物体,碳基生命利用了地球上丰富的碳资源和其他元素,通过一系列复杂的化学反应,形成了我们今天所见的多样生命形式。
那么,通过了哪些复杂的化学反应,形成了我们所见到的哪些生命形式呢?这还要从一个个具体的分子说起。
我们知道,构成生命的最基本分子是核酸和蛋白质,它们又是由一些基本的单元累积而成的,蛋白质的基本单元是氨基酸,核酸的基本单元是核糖核酸(RNA)和脱氧核糖核酸(DNA),科学家通过实验室模拟早期地球条件的实验,确定了氨基酸可以在地球上形成。同时还有研究表明,氨基酸可能是外太空的产物。也就是说,地球上有大量的蛋白质的基本单元。
对于核苷酸,现在也有大量的实验表明,嘌呤碱或嘧啶碱、核糖或脱氧核糖以及磷酸这些组成部分在地球早期环境中可能存在。海相起源说提出,在原始海洋中,小分子氨基酸和核苷酸可以吸附在黏土等物质的活性表面,在适当的缩合剂存在时,可以缩合成高分子聚合物,继而产生团聚体和原始细胞。
我们假设RNA或者DNA和蛋白质是在原始海洋产生之前就在那些著名的土著地点形成了,当大雨来临之时,他们被冲刷进地表的凹陷或者裂缝了,就在原地简单代谢的催化下,行成了相互融合的合作体,他们互相促进、互相支持,合作的非常愉快,从而数量日渐增多。
这时候,从外界漂浮而来很多脂肪分子,因为脂肪的密度比较低,他们漂浮在水面上,同时他们的作用也非常巨大,能够为RNA和DNA提供蔽体之衣,也就是让RNA和DNA有了区隔外界的膜,但是根据密度原理,ρ蛋白质 >ρ糖类 >ρ水> ρ脂肪。所以脂肪是漂浮在水面之上的,而在水底下的核酸、类核酸已经产生了很多,它们满溢在四周,竞争追逐着脂质双层膜囊,就像精子追逐着卵子,那些结合着蛋白质被保护的比较好的核酸率先闯进了脂质双层膜囊,没有进入到脂质双层结构的物质,就像是编外的,变成了病毒、类病毒、支原体、衣原体、立克次体或者散在的各种RNA,像魔法一样飘荡在四周的水域中。
说一下我们的基本结论:
1.DNA、RNA及氨基酸、蛋白质起源的比较早,在原始海洋之前就通过干湿循环形成了。
2.那场千年大雨,将它们冲刷到各种含能量的坑洼之地,造成了它们的复制繁荣。
3.简单的代谢需要水,在冲刷进RNA和DNA、蛋白质的坑洼之地进行着。
4.因为质膜密度比较小,当RNA和DNA、蛋白质比较多时,它们满溢四周,竞争结合者多种类型的膜成分。
5.没有进入到脂质双层结构的物质,就像是编外的成分,变成了病毒、类病毒、支原体、衣原体、立克次体或者散在的各种RNA,只能进行寄生、腐生和共生。
所以我们称为云雨起源说、冲洗(刷)理论,起源土著说、病毒编外说以及能量形式决定生命形式等等新说,既然假说已经提出,那么接下来我们我们需要的就是证明。
第五章 最简单的复制单元、代谢单元和膜脂(古菌单层膜)
以下七万字文字将陆续上传。




 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国