

全球的海洋生态系统现在正面临不少挑战,比如气候变化、海洋污染、过度捕捞,以及人类活动导致的栖息地破坏。这些问题让很多海洋生物,尤其是鲸类,面临生存危机,迫切需要我们关注和保护它们的栖息地和种群健康。南海是一个生物多样性丰富的地区,但在鲸类的栖息地和生态需求方面的研究还很不足,这让我们对它们的情况了解得不够多。这种知识的缺乏也使得我们很难制定出有效的保护措施。因此,科学家们认识到有必要填补这些空白,以便为未来的保护和管理工作提供所需的数据支持。目前,虽然国际上已经有了一些海洋保护的框架,但是,具体针对鲸类及其栖息地的政策和管理措施,还很不够。所以,中国科学家们希望通过这项研究,提供重要的数据,推动更好的政策制定和管理实践,从而保护这些脆弱的物种和它们的栖息地,并且助力《昆明-蒙特利尔全球生物多样性框架》(GBF)的目标实现。
“海洋与湿地”(OceanWetlands)小编注意到,2024年10月8日,来自中国科学院深海科学与工程研究所的李松海研究团队在《生物保护》(Biological Conservation)期刊上发表了一项重要研究,揭示了南海离岸及深潜鲸类的物种多样性及其关键栖息地。为助力全球环境治理、并供我国学者了解最新研究动态信息,编译分享信息如下,供感兴趣的读者们参阅。
这项研究指出,尽管南海被视为全球海洋生物多样性的热点区域,但在鲸类研究方面却存在严重的数据不足。这一现象在离岸鲸类物种的科学知识和保护基线信息方面尤为明显。为了填补这一空白,研究团队在2019~2023年间,进行了六次系统性的船舶调查,重点关注西沙-中沙群岛周边的鲸类分布。
在调查过程中,研究人员记录了211次鲸类目击事件,其中包括187次单一物种目击和24次混合物种目击。最终,他们识别出了17种齿鲸(odontocete),所有这些物种均为离岸及深潜型鲸类。研究发现,热带斑海豚(pantropical spotted dolphin)、短肢领航鲸(short-finned pilot whale)和弗氏海豚(Fraser's dolphin)是该地区的主要物种,因其频繁出现且数量相对丰富而被特别指出。

上图:南海海域的抹香鲸 中国科学院深海所海洋哺乳动物研究团队供图
基于本次研究的现场目击记录以及其它机会性目击数据,该研究团队确认:在南海栖息着至少25个鲸类物种。此外,该研究还结合了搁浅记录和标本,建立了一个包含至少36种鲸类的物种清单。这些发现不仅丰富了南海的生物多样性档案,也为未来的保护工作提供了重要的科学依据。
在生物地理学方面,研究显示,南海(尤其是西沙-中沙水域)的北部不仅支持高水平的鲸类物种多样性,还可能作为“易危”的抹香鲸(sperm whale)的育幼和觅食场所,并为许多其它离岸及深潜鲸类提供关键栖息地。为此,研究团队建议将西沙-中沙水域提名为重要海洋哺乳动物区域(Important Marine Mammal Area,IMMA),以便为这一重要区域的保护和管理提供支持。
该研究不仅填补了南海鲸类研究的空白,也引发了对保护这些珍稀海洋生物及其栖息环境的广泛关注。随着人类活动对海洋生态的影响日益加剧,保护措施的制定与实施显得尤为重要。该研究团队期望,通过这一研究,能够提高公众和相关部门对南海鲸类保护的重视,推动更有效的保护措施,确保这些海洋生物的持续生存与繁衍。

上图:一头抹香鲸。根据资料,抹香鲸可以在水底闭气超过一小时。©摄影:王敏幹(John MK Wong) | 绿会融媒·“海洋与湿地”(图文无关)
【引言】南海鲸类研究:保护海洋生态的重要指示器
鲸类作为海洋环境中的顶级掠食者,具有极高的生态重要性,能够反映海洋生态系统的健康状况(Rupil et al., 2022; Gilbert et al., 2023)。虽然现存的鲸类物种约有90种,仅占所有海洋生物(超过120万种)中的一小部分,但它们在维持海洋生物多样性方面发挥着重要作用(Pimiento et al., 2020; Kiszka et al., 2022)。近年来,鲸类保护已成为全球海洋保护倡议中的优先事项(Nelms et al., 2021; Alves et al., 2022; Mouton et al., 2022)。许多鲸类物种被认定为关键的保护对象,例如旗舰物种和伞护种,通常用于海洋保护区、国家公园、避难所和保护区的建立(Hoyt, 2012, 2022; Corrigan et al., 2014; Notarbartolo di Sciara et al., 2016)。
为了有效实施鲸类保护,获取有关鲸类物种多样性(Gill et al., 2015; Sankalpa et al., 2021)、分布范围(Correia et al., 2015; Di Tullio et al., 2016)、种群丰度(Hammond et al., 2013; Rogan et al., 2017)、关键栖息地(如觅食和育幼场所;Liu et al., 2022a; Zhang et al., 2023)及其生存威胁(Campana et al., 2022; Braulik et al., 2023)的可靠科学数据至关重要。开展系统、设计严谨的鲸类专用调查是获取这些数据的最直接和基础的方法,通常通过船舶或航空器进行(Eberhardt et al., 1979; Kinzey et al., 2000; Williams and Thomas, 2009)。然而,在许多发展中国家和地区,这种田野调查方法的使用严重不足,主要归因于科学关注的不足和资金支持的匮乏(Kaschner et al., 2012; Tyne et al., 2016; Astudillo-Scalia and de Albuquerque, 2020)。因此,在一些具有高海洋生物多样性的热点水域(如南海)中,鲸类专用的科学调查努力通常显得不足(Tittensor et al., 2010; Kaschner et al., 2012)。
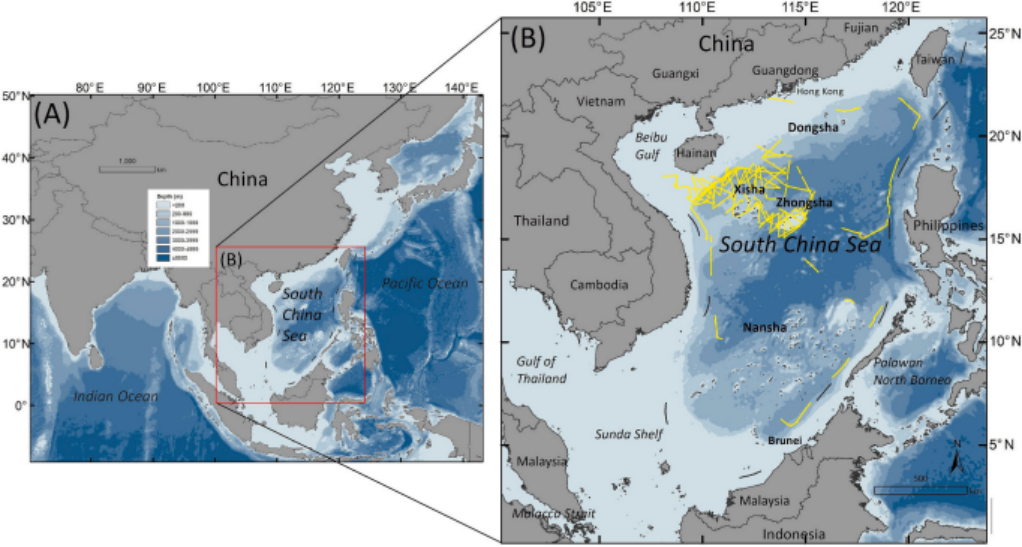
图1. (A)印度-太平洋区域(包括西太平洋和东印度洋)地图;(B)南海区域局部放大图。图中黄色线条表示2019年至2023年间开展的针对鲸豚类动物的专项调查航线。(有关图例中颜色表示的详细信息,请参阅本文的网络版。)图片来源:Liu M, Lin W, Lin M, et al.
南海的面积约为350万平方公里(1°N - 22°N,104°E - 122°E),是热带亚热带地区最大的边缘海(Morton and Blackmore, 2001)。该海域被广泛认定为全球生物多样性热点,位于全球海洋生物多样性中心(即珊瑚三角区)的西界,展示了极高的海洋物种丰富度(Kaschner et al., 2011; Jenkins and Van Houtan, 2016; Astudillo-Scalia and de Albuquerque, 2020)。南海作为一个半封闭海域,周边环绕着中国、越南、菲律宾、马来西亚和印度尼西亚等多个大型发展中国家(图1B)。这些国家的沿海亚区居住着超过7亿人,占全球人口的约9%(联合国经济和社会事务部,2024年)。近年来,南海面临着多种强烈且持续的人为压力,例如渔业、化学和垃圾污染、海洋工程建设和航运。因此,许多濒危物种,如鲸类、珊瑚礁、海龟以及鲨鱼和鳐鱼的保护问题引起了越来越多的关注。
根据历史捕鲸和搁浅信息,南海已记录超过三分之一的现存鲸类物种(即全球90种鲸类中的30种以上)(Wang, 2012; Liu et al., 2022b),这表明,该区域具有较高的鲸类物种丰富度。但是,目前针对这些物种的可靠科学数据仍然匮乏,制约了该区域的海洋保护工作(Lin et al., 2019; Li et al., 2020)。值得注意的是,现有的鲸类研究大多集中于近岸物种,例如印太驼海豚(Sousa chinensis; 又称中华白海豚)、伊拉瓦底海豚(Orcaella brevirostris)和江豚(Neophocaena spp.)(Perrin, 2002; Lin et al., 2019; Li, 2020)。其主要原因在于,近岸物种相较于离岸鲸类更易接触,减少了沿海调查的成本(Aragones et al., 1997; Kaschner et al., 2012; Tyne et al., 2016)。到目前为止,针对南海离岸鲸类的知识基础主要限于偶然搁浅(Vu and Ponnampalam, 2018; Liu et al., 2019; Li et al., 2021; Chou et al., 2024)、误捕事件(Zhao et al., 2017; Liu et al., 2017; Lin et al., 2023)、标本(Yamada and Chou, 2004; McGowen et al., 2021)和机会性目击(Smith et al., 1997, 2003; Yang et al., 1999; De Boer, 2000; Ponnampalam, 2012; Hu et al., 2023)。尽管这些零散数据可以提供鲸类物种存在及区域丰富度的初步指示,但无法为该区域迫切的鲸类保护需求提供可靠的基线信息。
为了填补这一知识空白,研究团队在南海开展了多次专门针对鲸类的船舶和直线样线调查,主要覆盖离岸及深海水域。调查中,研究人员对可能出现在研究区域的所有鲸类物种进行了详细调查。基于现场数据,他们提供了关于离岸及深潜鲸类的宝贵保护信息,包括分布模式、物种丰富度、相对丰度及栖息地特征,这些信息对该区域的鲸类保护管理策略至关重要。结合其它信息来源,如机会性目击、搁浅记录和标本,该研究团队进一步分析了南海不同海区鲸类的区域物种丰富度及生物地理模式。总体而言,研究目标是为南海离岸和深潜鲸类的有效保护措施制定提供科学依据,并为未来的鲸类保护研究指明方向。

上图:科研人员在南海记录到短肢领航鲸与弗氏海豚混游现象。图片来自新华社
【研究方法与材料】南海海洋哺乳动物调查
2.1 调查区域
南海(SCS)是一个环境多样的区域,包括岛屿和礁石周围的浅水区、大陆架区域(深度≤200米)及深水区(深度:200-5377米)(Morton and Blackmore, 2001)。自2019年至2023年间,该研究团队一共完成了六次调查。在前四次调查中,他们选择了位于南海西北部的西沙群岛和中沙环礁周围的深水区(以下简称“西沙-中沙水域”)作为主要的船舶调查区域(Lin et al., 2021; Rosso et al., 2021; Liu et al., 2022a)。第一次调查的初步结果已由Lin等人(2021)和Rosso等人(2021)报告。在第五次调查中,研究团队首次对越南中南部及南沙群岛周边水域进行了考察(图2E)。第六次调查则主要集中在菲律宾西部水域及东沙群岛(图2F)。所有调查的水域都呈现出多样的海底地貌,包括大陆架边缘、陡峭的大陆坡、海山和海底峡谷(Wang et al., 2018; Ou et al., 2021)。以往的研究表明,这些海底地貌可能是短肢领航鲸(Globicephala macrorhynchus)、抹香鲸(Physeter macrocephalus)和各种喙鲸(Ziphiidae)等多种海洋哺乳动物偏爱的栖息地(Davis et al., 2002; Gill et al., 2015; Breen et al., 2020; Correia et al., 2021)。
2.2 调查设计
在船舶调查期间,研究团队使用了一艘长44米、吨位500吨的研究船,以约7-8节的巡航速度(即13-15公里/小时)进行专门的鲸类调查(Rosso et al., 2021; Liu et al., 2023a)。他们采用了线性横断面采样调查设计,通过之字形的航线覆盖海南岛附近的调查区域(Strindberg and Buckland, 2004; Thomas et al., 2007)。为了到达南海偏远的区域,调查团队根据海洋条件进行了机会性航线的调整。研究船的前顶层设有一个高度约7米的视觉观察平台,观察者可以在有效调查时间内向前方180度区域搜索鲸类,或在焦点跟踪期间观察周围360度的区域(Lin et al., 2021; Liu et al., 2022a, 2024a)。为了提高发现鲸类的概率,研究团队组建了两个观察组:主观察组(POs)和独立观察组(IOs),两组同时工作(Eberhardt et al., 1979; Kinzey et al., 2000)。在视觉观察平台上设置了五个值班位置:PO组的左、中、右,IO组的左和右。PO组由5~8名经过良好训练的观察员组成,他们在每个PO位置上轮班,每半小时更换一次,使用7×50手持双筒望远镜(Fujinon FMT-SX或Burris Caribbean)。IO组则由3~4名经验丰富的高级研究人员组成,他们从左到右轮流担任各自的IO位置,每半小时更换一次,使用7×50手持双筒望远镜或Fujifilm 20×150 Bigeyes双筒望远镜。根据上述值班周期,所有参与观察的人员在两个值班观察期之间至少休息一小时,以减少视觉疲劳。调查主要在每年的四月至九月进行,这一时期西南季风对南海的海洋条件有显著影响(Liu et al., 2023a, 2024a)。此外,调查仅在良好的视觉条件下进行(东八区白天;0800-1800时无雾或降雨),当海洋条件不佳(如博福特风级≥5)时,研究团队会暂停寻找鲸类,寻找更合适的区域进行调查。
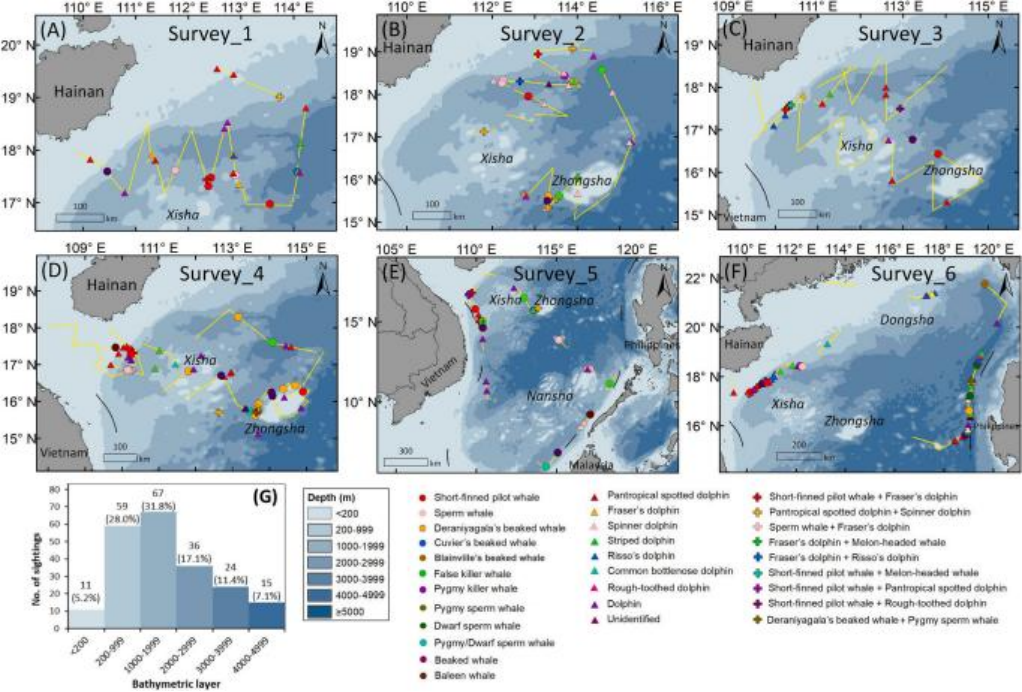
(上图2. (A-F)南海记录到的鲸豚类物种的目击地点和分布格局。A-F图分别对应表1中的第1-6次调查。(G)所有目击地点的海水深度直方图。图片来源:Liu M, Lin W, Lin M, et al. )
2.3 数据收集
一旦遇到鲸类,研究船的巡航速度将减至3-5节(约5-10公里/小时)以接近鲸群(Lin et al., 2021; Liu et al., 2022a)。在跟踪鲸类时,会暂停在轨道上的视觉观察,直到跟踪结束后再恢复观察(Liu et al., 2023a, 2024a)。每次观察时,两名在职的IO负责收集遇到动物的信息,包括日期、时间、GPS位置、物种、与船只的距离、动物群体与航行方向的角度、行为、群体大小(低估/最佳估计/高估)、母仔对的存在/数量及其它附加描述(Kinzey et al., 2000; Correia et al., 2015; Di Tullio et al., 2016)。在中间位置的PO负责记录所有详细信息,并使用标准化的数据表进行记录。尽可能情况下,值班IO会在物种水平上识别每次观察(Arcangeli et al., 2017; Bilgmann et al., 2018)。然而,对于某些观察,由于现场的物种鉴定只能达到较宽泛的分类水平,如无法识别的海豚(Delphinidae)、喙鲸(Ziphiidae)、齿鲸(Odontoceti)或须鲸(Mysticeti)(Di Tullio et al., 2016; Braulik et al., 2018)。
当实验条件允许时,研究团队会从母船(即44米长的研究船)后部部署一艘小型刚性充气船(RHIB;长8.8米,配备150 HP四冲程发动机)接近遇到的动物。通常情况下,至少有2~3名经验丰富的摄影师负责使用数码相机、无人机和水下相机拍摄遇到动物的高质量照片和视频(Liu et al., 2021, 2022a, 2024a)。遇到动物的水下声音则使用自主声学记录仪进行录制(Dong et al., 2023, 2024a, 2024b)。一旦遇到动物靠近时,研究团队将从母船和RHIB收集照片和水下声音(Dong et al., 2024a, 2024b; Liu et al., 2024b)。在RHIB上,团队还使用Paxarms MK24C喷射器进行一些隐蔽物种的活体组织样本采集。对组织样本可以进行遗传和基因组分析(Lin et al., 2024)。总体而言,这些影像、生物声学和遗传数据对于在后续调查中加强物种鉴定尤为重要,能够修正、补充或甚至交叉验证物种鉴定结果(Rosso et al., 2021; Dong et al., 2024a, 2024b; Liu et al., 2024b)。
2.4 数据分析
研究团队使用EstimateS 9.0版绘制物种鉴定累积曲线,以判断其调查工作是否充分代表了该区域的物种丰富度(Colwell, 2013; Di Tullio et al., 2016)。每次调查的航线和鲸类观察位置均通过ArcGIS 10.1版(ESRI,Redlands, CA, USA)进行展示。此外,他们还绘制了所有鲸类观察位置的深度值直方图,以展示水下特征。每种物种的常见程度或稀有程度则通过出现的调查次数(范围1至6次)进行确定。由于空间覆盖不均(图1B)以及某些物种观察次数较少,无法估算记录物种的绝对丰度(例如种群大小)(Liu et al., 2022a, 2024a)。相反,研究团队计算了两个参数来表示各物种的相对丰度:群体观察率(GSR;每100公里调查工作中遇到的群体数量)和个体观察率(ISR;每100公里调查工作中遇到的个体数量)(Hammond et al., 2013; Sankalpa et al., 2021; Giralt Paradell et al., 2023)。
此外,他们还利用R 4.0.5版(R开发核心团队,2021)中的“heatmap.2”包绘制了观察率(GSR和ISR)和物种常见/稀有程度的热图矩阵。为了减少潜在偏差,未包括夜间和声学探测等离线观察(Hamilton et al., 2021; Filatova et al., 2022)。在去除所有焦点跟踪期间的观察后,研究团队使用ArcGIS计算了所有六次调查的在横断线调查工作强度(视觉搜索工作),以0.5°×0.5°的网格图展示(Sankalpa et al., 2021; Liu et al., 2022c)。
2.5 信息整合
为了补充实地观察的数据集,研究团队构建了一个基于文献、社交媒体、机会性航行和个人沟通所收集的海南岛及周边海域的机会性鲸类观察数据集(见表S1)。为了提供南海鲸类物种的全面清单,研究团队进一步审查了文献、搁浅报告及其它来源的所有鲸类发生记录,采用与Liu等人(2023b, 2024a)相同的方法。各种数据来源的详细信息,如观察记录、搁浅数据库和标本/骨骼标本收藏,见表S2。尽管这些数据源大多仅覆盖南海的小区域或沿海地区,但通过区域尺度的数据整合,依然可以获得具有可比性和保护价值的信息。利用这些数据集,研究团队量化了两个关键的生物多样性指标:
鲸类物种丰富度(即记录的鲸类物种数量)和优势物种(即所有记录中前三个物种所占的比例)(Braulik et al., 2018; Liu et al., 2022b)。研究团队在南海地图上绘制了这两个指标,以展示不同生态区的鲸类群落“推断”生物地理模式(Correia et al., 2020; Purdon et al., 2020)。南海不同生态区的划分依据是Spalding等人(2007)的研究。

江豚。©摄影:王敏幹 | 绿会融媒·“海洋与湿地”(图文无关)
【研究结果】
3.1 调查努力与物种丰富度
从2019年4月~2023年7月,研究团队在调查区域进行了六次调查,共计120天的调查时间,累计调查距离达15,282公里(见表1)。每次调查的路线和鲸类观察位置如图2A–F所示。团队记录了211次鲸类观察,其中包括187次单一物种(占88.6%)和24次混合物种(占11.4%)的观察(见表1)。在所有记录的群体中,76.6%(180/235)可在物种水平上进行识别,而55条记录(23.4%)仅能在广义分类水平上进行确认,包括15组未识别的喙鲸(Ziphiidae)、2组未识别的须鲸(Mysticeti)、10组未识别的齿鲸(Odontoceti)和28组未识别的海豚(Delphinidae)(见表2)。
在每次调查中,研究团队至少识别了8种,最多识别了15种鲸类(见表1)。总共有17种齿鲸通过视觉观察、照片以及额外的遗传和声学证据被识别(见表2)。随着调查努力的增加,观察到的物种数量在第一至第20天之间迅速积累(即在第20天时记录了11种),在第21至第80天之间逐渐放缓(即在第60天时记录了15种),并在第100天后达到了17种的渐近值(见图3)。所有17种确认的物种在第一次到第四次调查中均成功识别。在第85天之后,即第五次和第六次调查中没有出现新的确认物种(见图3)。
3.2 物种普遍性、优势物种与相对丰度
热带斑点海豚(Stenella attenuata)是最具优势的物种,表现出最高的群体观察率(GSR,0.242组/每100公里)和个体观察率(ISR,9.174个体/每100公里)(见图4A和4B)。短肢领航鲸和弗氏海豚(Lagenodelphis hosei)分别位列第二和第三,具有相对较高的GSR(分别为0.151和0.124组/每100公里)和ISR(分别为7.499和6.576个体/每100公里)(见图4A和4B)。相比之下,其余14种物种的GSR(0.013–0.111组/每100公里)和ISR(0.013–4.103个体/每100公里)均低于前三种优势物种(见图4A和4B)。
短肢领航鲸、热带斑海豚、弗氏海豚、以及条纹海豚(Stenella coeruleoalba)是最常见的物种,这四种物种在所有六次调查中都经常被观察到(见图2A–F和图4A–B)。此外,抹香鲸和飞旋海豚(Stenella longirostris)也相对常见,分别在五次调查中记录(见图2A–F和图4A–B)。相比之下,柏氏中喙鲸(Mesoplodon densirostris)、小虎鲸(Feresa attenuata)、瓜头鲸(Peponocephala electra)、瓶鼻海豚(Tursiops truncatus)和糙齿海豚(Steno bredanensis)则表现出低普遍性,仅出现在两次调查中。
从空间分布来看,调查努力覆盖了154个网格单元(0.5° × 0.5°),主要集中在西沙-中沙水域(见图5A)。调查中确定了多个高观察密度的热点区域(见图5B)。混合物种的观察在海南南部、西沙-中沙和菲律宾西部的水域被发现(见图5C)。在调查的网格单元中,“黑鲸”(见图5D)和喙鲸(见图5F)的观察密度相似(分别为13.6%,21/154和12.3%,19/154);然而,它们在细尺度上的分布单元存在差异。抹香鲸(见图5E)和小抹香鲸属动物(Kogia spp.)(见图5G)的分布区域相对有限(分别为6.5%,10/154和3.9%,6/154)。尤其是,海洋海豚(见图5H)具有最广泛的分布区域,占调查网格单元总数的45.5%(70/154)。
3.3 社会特征
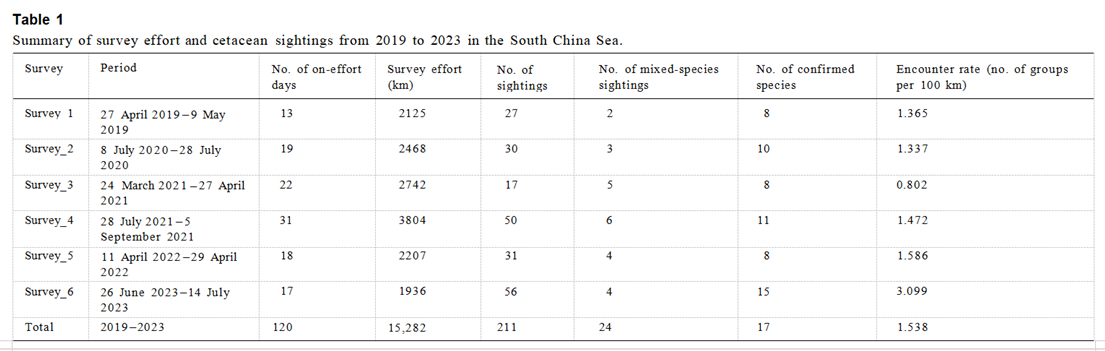
(上图:表1:2019-2023年南海鲸豚类调查情况汇总。)在记录的群体中,42.1%(99/235)被识别为母性群体,确认存在幼体如幼鲸和/或未成年(见表2)。多个物种表现出较高的母性群体数量及比例,包括短肢领航鲸(78.2%;18/23)、抹香鲸(46.7%;7/15)、热带斑海豚(50.0%;19/38)、弗氏海豚(61.9%;13/21)、飞旋海豚(56.3%;9/16)、条纹海豚(62.5%;10/16)及瑞氏海豚(Grampus griseus;83.3%;10/12)(见表2)。
所有24个混合物种群体均涉及两个相关物种。记录的大多数物种(85.6%;11/17)可能包括混合物种群体,并观察到10对物种对(见表S3)。在所有记录中,“弗氏海豚与短肢领航鲸”的组合是最常见的,记录了九次观察(37.5%;9/24)。此外,“热带斑海豚与飞旋海豚”的组合也相当常见,记录了四次观察(16.7%;4/24)。
在样本量达到10个以上的八种物种中,群体规模存在差异(Kruskal-Wallis检验,KW = 65.529,df = 7,p < 0.001)(见图6A和表3)。抹香鲸(4.9 ± 2.1;n = 15)和德拉尼亚加喙鲸(Mesoplodon hotaula;2.5 ± 0.8;n = 12)的平均群体规模最小,与短肢领航鲸(45.3 ± 41.7;n = 23)相比存在显著差异(p = 0.041和0.036)。短肢领航鲸(45.3 ± 41.7;n = 23)、热带斑海豚(45.2 ± 48.1;n = 38)、弗氏海豚(55.8 ± 36.4;n = 21)、飞旋海豚(37.8 ± 60.8;n = 16)和条纹海豚(36.9 ± 23.4;n = 16)则显示出相对较大的平均群体规模,而瑞氏海豚(16.8 ± 13.5;n = 12)则具有中等的平均群体规模(见图6A和表3)。
3.4 栖息地偏好
如图2A–F所示,大多数调查路线(91.2%;13,937公里/15,282公里)及绝大多数观察(94.8%;200/211)都发生在≥200米的深水区。在211次观察中,28.0%(59/211)和31.8%(67/211)发生在200–999米和1000–1999米的深水区;而只有17.1%(36/211)、11.4%(24/211)和7.1%(15/211)发生在2000–2999米、3000–3999米和4000–4999米的深水区。一些栖息地特征在样本量为10个以上的八种物种之间存在差异,而另一些则表现出相似性。Kruskal-Wallis检验显示,物种间在水深(KW = 35.183,df = 7,p < 0.001)和距200米的距离(dist_200)(KW = 14.992,df = 7,p = 0.036)上存在显著差异;而在坡度(KW = 9.440,df = 7,p = 0.223)和距1000米的距离(KW = 13.376,df = 7,p = 0.062)上没有显著差异(见图6B和表3)。具体而言,热带斑海豚、短肢领航鲸、弗氏海豚和飞旋海豚显著偏好较浅的栖息地(p < 0.05),而抹香鲸、瑞氏海豚、梅伦头鲸和侏儒杀人鲸则显著偏好较深的栖息地(p < 0.05,见图6B和表3)。
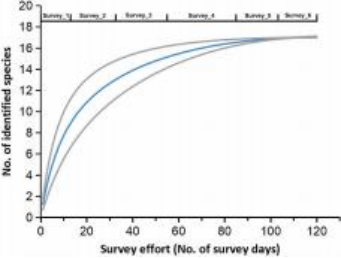
图3. 南海六次调查中鲸类物种丰富度的累积曲线。X轴表示调查努力量,即调查天数;Y轴表示鉴定出的鲸类物种数。图源:Liu M, Lin W, Lin M, et al.
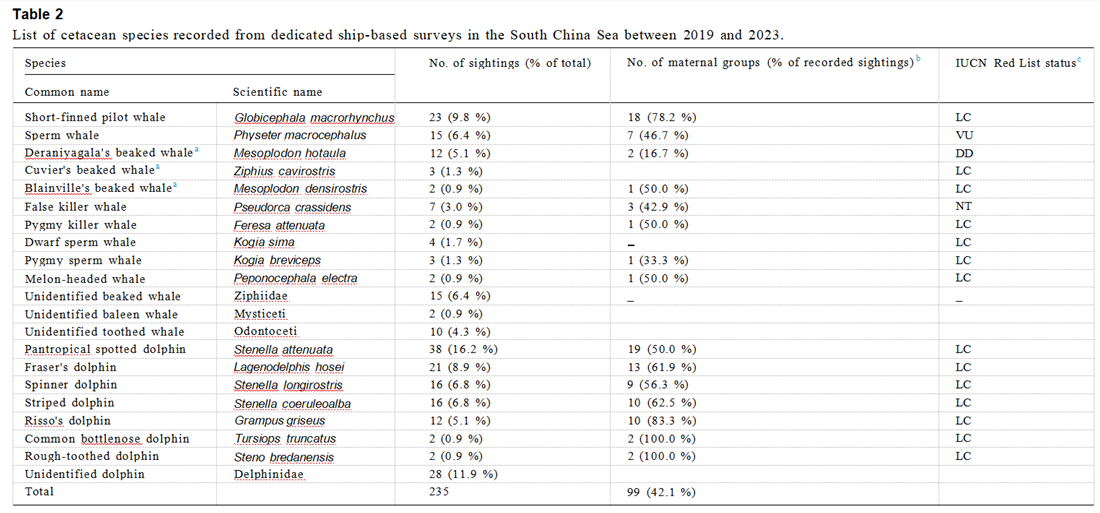
表2:2019年至2023年期间,在南海进行的专项船上调查记录的鲸豚物种清单。图源:Liu M, Lin W, Lin M, et al.

图4. 南海六次调查中鲸豚类相对丰度的比较。(A)群体目击率(GSR,每100公里调查努力量所遇到的群体数);(B)个体目击率(ISR,每100公里调查努力量所遇到的个体数)。物种的常见程度由其在几次调查中出现来表示(范围为1至6)。图源:Liu M, Lin W, Lin M, et al.
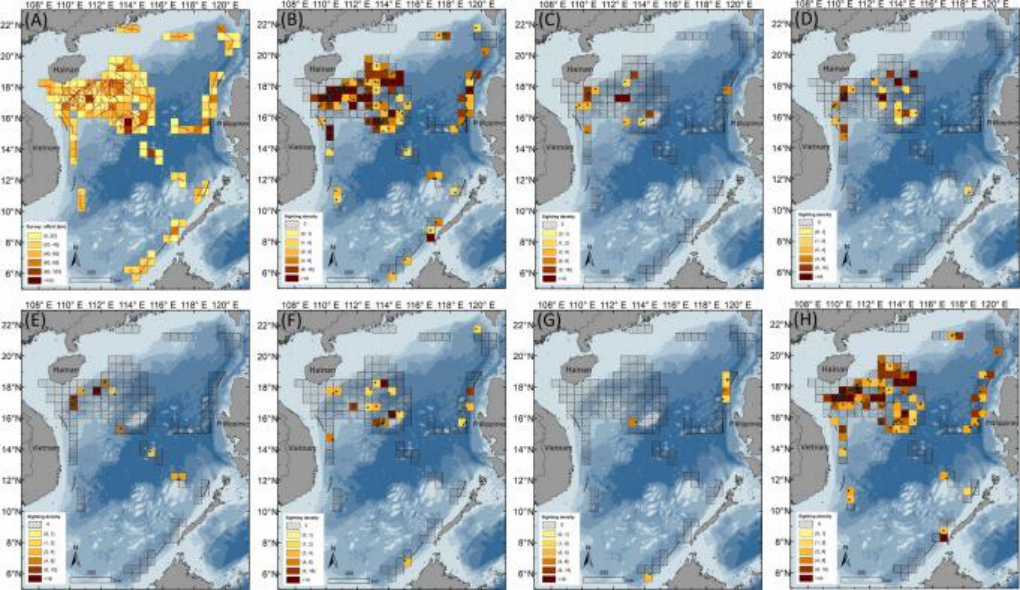
图5. (A)0.5°×0.5°网格单元图显示的调查努力强度。鲸豚类目击密度(即每100公里调查努力量所遇到的鲸豚类目击次数)显示为:(B)所有鲸豚类目击(n=211),(C)混合物种组合(n=24),(D)“黑鱼”包括短肢领航鲸、小虎鲸、伪虎鲸和瓜头鲸(n=34),(E)抹香鲸(n=15),(F)喙鲸物种包括德氏中喙鲸、柯氏喙鲸、柏氏中喙鲸和其它未识别的喙鲸(n=32),(G)侏儒抹香鲸和小抹香鲸Kogia spp.(n=7),以及(H)远洋海豚,包括热带斑海豚、弗氏海豚、飞旋海豚、条纹海豚、瑞氏海豚、瓶鼻海豚、糙齿海豚和其它未识别的海豚(n=135)。图源:Liu M, Lin W, Lin M, et al.
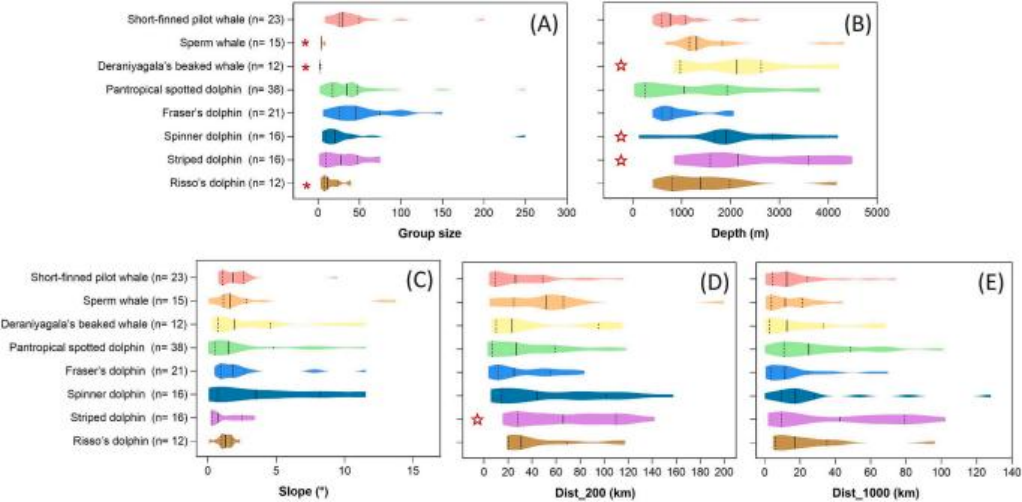
图6. 南海调查区域内目击次数大于10次的八种鲸豚类物种的群体大小和栖息地特征。小提琴图为(A)群体大小(即每个群体中估计的个体数量),(B)深度(即每个目击地点的水深;米),(C)坡度(度),(D)dist_200和(E)dist_1000(即每个目击地点与200米和1000米等深线的距离;公里)。图中显示了中位数(实线)、25%和75%四分位数(虚线)。星号(*)表示与短肢领航鲸相比显著较低的物种,星号(☆)表示与短肢领航鲸相比显著较高的物种。图源:Liu M, Lin W, Lin M, et al.
【讨论】
物种组成、优势物种与分布
研究团队的实地观察数据表明,在调查区域内,热带斑海豚、短肢领航鲸和弗氏海豚是三种主要的优势物种(见图4A–B),它们主要分布在西沙-中沙水域(见图2A–F和5B)。这一发现的原因在于这些物种具有较高的常见度、较高的观察频率、较大的相对丰度以及较大的群体规模和深水栖息地特征(见图6B–E)。这一结果与海南岛周边的机会性鲸类观察数据中发现的优势物种有所不同(见表S4),后者主要记录了生活在近岸水域的中华白海豚和布氏鲸(Liu et al., 2022c, 2023c; Zhang et al., 2023)。然而,这一结果与海南地区的搁浅记录部分一致,热带斑海豚是海南岛周边最常见的搁浅物种之一(Liu et al., 2019, 2022b)。相比之下,短肢领航鲸和弗氏海豚在海南的搁浅率显著低于包括印太江豚(Neophocaena phocaenoides)和布氏鲸在内的近岸物种(Liu et al., 2019, 2022b)。这种差异可能是因为搁浅记录的频率受到物种栖息地偏好的影响(例如水深和离岸距离)及区域丰度的影响(Thompson et al., 2013; Olson et al., 2020)。
从空间分布上看,研究团队的实地观察数据强调,海洋海豚(包括热带斑海豚、弗氏海豚、飞旋海豚、条纹海豚、瑞氏海豚、瓶鼻海豚和糙齿海豚)在南海的调查区域广泛分布(见图5H)。这些海洋海豚的分布模式通常与其已知生态学相一致,显示出在温暖的热带和温暖温带海域有广泛的分布范围(Jefferson et al., 2015; Mannocci et al., 2020)。此外,研究中涉及到的海洋海豚至少有七个物种,其物种丰富度高于其它分类群(见表2)。与此相反,“黑鲸”物种(包括短肢领航鲸、小虎鲸、伪虎鲸和瓜头鲸)、抹香鲸、喙鲸科(德氏中喙鲸、柯氏喙鲸和柏氏中喙鲸)以及小抹香鲸属(侏儒/小抹香鲸)显示出较小的活动范围和相对受限的栖息地特征(见图5D–G和6B–E)。
除了专门调查收集的实地观察数据,研究团队的补充数据进一步证实,热带斑海豚和短肢领航鲸是南海的两种常见物种。这两种物种在中国的海南、广西、广东和台湾省是常见的搁浅物种(Liu et al., 2019, 2022b; Li et al., 2021; Chou et al., 2024),同时也在越南海岸(Vu和Ponnampalam, 2018)出现,仅次于其它近岸齿鲸物种,如中华白海豚和江豚。从更广泛的角度来看,多个研究报告表明热带斑海豚在南海的许多区域,包括中国台湾沿海(Yang et al., 1999; Wang et al., 2001; Hu et al., 2023)、海南(Gong et al., 2019)、东沙群岛(Fu et al., 2022)、越南(Smith et al., 2003)、菲律宾的吕宋和巴拉湾(De Boer, 2000; Aragones et al., 2024)以及马来西亚的砂拉越(Ponnampalam, 2012)均有记录。同样,关于“黑鲸”物种(尤其是短肢领航鲸)和抹香鲸的机会性观察和搁浅数据也揭示了它们在南海深水区的广泛分布模式(De Boer, 2000; Moore和Lien, 2007; Tseng et al., 2011; Ponnampalam, 2012; Hu et al., 2023; Liu et al., 2023b, 2024a)。这些补充证据表明,当前的调查区域并未覆盖南海深潜鲸类的所有潜在适宜栖息地,研究团队推测这些物种可能在南海的广阔深水区域中广泛分布。
研究团队在第五次、和第六次调查中收集的实地观察数据(见图2E–F)为越南中南部(岘港和广义)及菲律宾西部(吕宋和巴拉湾)水域的鲸类物种的出现和组成提供了一些见解。结果显示这些区域的物种出现和组成与越南的搁浅和骨骼标本记录存在一些相似性(Vu和Ponnampalam, 2018; McGowen et al., 2021)以及菲律宾西部的搁浅记录(Aragones et al., 2024)。更重要的是,研究团队的实地观察证实了这些区域内几种难以观察的物种的存在,包括德氏中喙鲸、柯氏喙鲸和柏氏中喙鲸(Dong et al., 2023, 2024a; Liu et al., 2024b),这些物种在标本或搁浅记录中并未得到充分揭示(Vu和Ponnampalam, 2018; McGowen et al., 2021; Liu et al., 2022b; Aragones et al., 2024),或在机会性观察中(Smith et al., 1997, 2003; Yang et al., 1999; De Boer, 2000; Ponnampalam, 2012)。这些物种的遗漏表明,虽然其它信息来源可以补充一些证据,但实地调查对于获取鲸类物种出现和分布的基础信息至关重要。
社会行为特征与关键栖息地
研究团队的实地观察数据显示,南海的鲸类展示了复杂的种内及种间社会行为特征,频繁观察到母子群体(见表2)、多个海豚物种的较大群体规模(见图6A)以及多样的混合种群体对(见表S3)。不仅在小型海豚物种(如热带斑海豚、弗氏海豚、飞旋海豚、条纹海豚和瑞氏海豚)中,研究团队还观察到中型至大型齿鲸物种(如短肢领航鲸和抹香鲸)中也频繁出现母子群体(见表2)。结合研究团队确定的深海栖息地偏好(见图6B–E和表3),这些结果表明,研究团队调查的南海区域,特别是西沙-中沙水域,可能是许多鲸类物种关键生存周期活动的重要栖息地(Lin et al., 2021; Rosso et al., 2021; Liu et al., 2024a)。特别是,西沙-中沙水域被假设为西太平洋抹香鲸种群的育儿和/或觅食场(Liu et al., 2022a, 2023b)。
与抹香鲸和喙鲸不同,观察到的南海小型海豚通常形成相对较大的群体(见图6A)。这可能是因为生活在大群体中是海豚社会中一种普遍的社会和生存策略(Gowans et al., 2007; Mesnick et al., 2019)。此外,混合种群体中可能也会获得社群生活的好处(见表S3),特别是在它们具有相似的需求和能力时,例如在同一区域生活、以相似的猎物为食、以相似的速度游泳、抵御相同的捕食者并共享声学社交信号(Qurouil et al., 2008; Kiszka et al., 2011; Syme et al., 2021)。在调查区域内,短肢领航鲸与弗氏海豚的混合种群体被观察到,这可能指示着某种社会关系(见表S3),尽管在现有文献中,关于两者之间的相互作用的记录相对较少(Owen et al., 2023)。此外,这些社交群体结构的形成和演变仍有待未来研究的深入探讨。
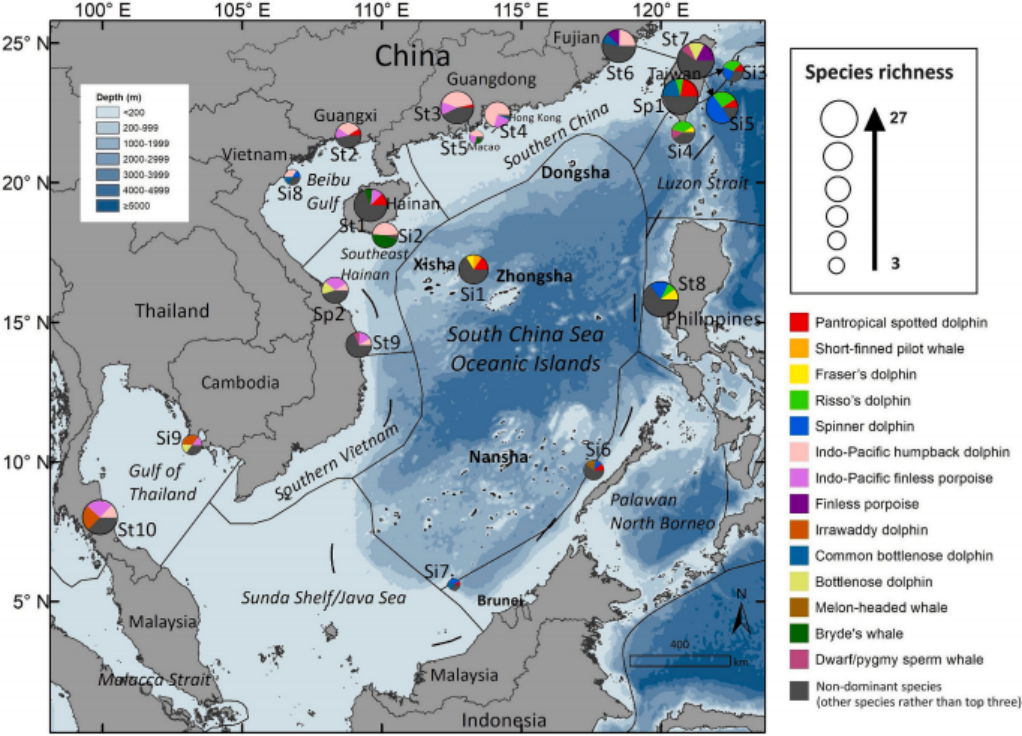
图7. 基于不同数据来源确定的南海各海域鲸豚类物种丰富度(饼图大小代表记录物种数量)和优势种(饼图中前三种记录物种所占比例)。子海域的划分依据Spalding等人(2007)的研究。关于数据来源的详细信息,即目击记录、搁浅数据库和标本收集,请参见补充表S2。每个饼图的位置代表不同数据来源的集中调查区域或覆盖范围。Liu M, Lin W, Lin M, et al.
【结论与展望】
综合这项研究的发现,调查结果为南海鲸类物种的出现、生态和分布模式提供了新的认识和见解。尤其是在西沙-中沙水域发现的多个重要物种,不仅对保护工作具有重要意义,还为区域海洋生态系统的完整性提供了支撑。尽管已经取得了诸多发现,但南海的深水区域仍然需要更广泛和深入的实地调查。研究团队建议定期开展现场监测和科学研究,以为海洋保护和管理政策提供信息,以更好地理解和保护南海及其生物多样性,为可持续的海洋环境和生态系统恢复提供数据支持。
【研究数据的附加信息】
本次研究使用的部分数据来自多位专家学者提供的文献及资料。此外,研究团队重申,在调查过程中,对鲸类的观察数据进行了细致的记录,并与现有数据进行了比较,以确保对研究结果的准确性。希望本研究的结果能为今后的相关研究提供参考,推动南海及其它区域鲸类生物学及其保护工作的持续深入。
海湿·小百科
01关键栖息地(Critical Habitat)是指对于某些特定物种的生存、繁衍和生长至关重要的自然栖息环境。这些栖息地通常提供物种所需的食物、庇护和繁殖场所,因此保护关键栖息地是维持生物多样性和生态平衡的重要措施。关键栖息地的定义通常基于物种的生态需求和生存威胁,确保这些区域得到合理的管理和保护,对于防止物种灭绝和促进生态恢复至关重要。
在本文中,之所以提到“关键栖息地”这个概念,是为了强调某些特定区域对于海洋哺乳动物的重要性,比如海豚和鲸。研究团队通过观察发现,西沙-中沙的水域为这些动物提供了很好的栖息环境,特别是它们的育幼和觅食场所。明确这些区域作为关键栖息地,有助于科学家、政策制定者和保护组织制定保护措施,确保这些地方的生态功能得以保持,从而促进海洋生物的可持续发展和保护。所以说,识别和保护这些关键栖息地是维持海洋生态系统健康和保护生物多样性的一个重要环节。
**02物种组成
物种组成**(Species Composition)是指在特定生态系统或环境中,各种生物物种的种类及其相对丰度的组合情况。它不仅包括所观察到的物种种类,还涉及这些物种在群落中所占的比例和数量。通过分析物种组成,研究人员能够了解生态系统的多样性、物种之间的相互关系以及环境条件对物种分布的影响。在本研究中,对于“物种组成”的分析,能够帮助研究团队识别出在南海特定区域内的主要海洋哺乳动物种类及其分布模式,从而为保护策略的制定提供科学依据。
**03混合种群体
混合种群体**(Mixed Species Groups)是指在同一时间和空间内,由不同物种组成的群体。这种现象在许多生态系统中其实都很常见,特别是在海洋、森林等多样性较高的环境中。混合种群体的形成,往往跟物种间的相互关系有关,比如说,有相似的生活习性、共同的食物来源、共同的栖息地需求等。通过在混合种群体中生活,物种可能获得一定的生存优势,比如提高捕食者的警惕性、增加觅食效率,或是提高繁殖成功率。在上面的这个研究中,对“混合种群体”的观察,给该研究团队提供了关于不同物种如何共存以及它们在生态系统中相互作用的有价值信息。
**04重要海洋哺乳动物区域
重要海洋哺乳动物区域**(Important Marine Mammal Areas, IMMAs)是指在海洋生态系统中,对特定海洋哺乳动物种群具有重要意义的栖息地。这些区域被识别为具有保护潜力,能够为海洋哺乳动物提供繁殖、觅食及栖息等必要环境。IMMAs的设立旨在弥补传统海洋保护区在保护海洋哺乳动物方面的不足,确保这些脆弱物种在生态系统中的生存与健康。
通过采用科学标准和专家评估,IMMAs不仅有助于提升海洋哺乳动物的保护地位,还能够促进更广泛的生物多样性保护。它们帮助识别和划定对海洋哺乳动物生存至关重要的栖息地,支持全球和地区的海洋保护措施,如海洋保护区(MPAs)和生态或生物重要区域(EBSAs)的设立。而且,海洋哺乳动物作为指示物种,能够反映海洋生态系统的健康状况,IMMAs的实施将有助于更全面地维护和恢复海洋环境,确保海洋生物多样性和生态系统的健康。
IMMAs是由世界自然保护联盟(IUCN)物种存续委员会(SSC)与海洋哺乳动物保护区工作组(MMPATF)联合制定的标准进行认定的。IMMAs的选择标准,是在2013年法国马赛的研讨会上首次制定的,并经过了多次的修订、公众咨询。自2016年以来,已举办了多个地区专家研讨会来识别和确认这些区域。截至2021年10月,全球已识别出173个IMMAs,此外还有23个候选IMMAs和140个关注区域(Areas of Interest),这标志着对海洋哺乳动物保护的重要进展。
“海洋与湿地”(OceanWetlands)小编注意到最新的数据是:截至2024年,全球范围内已经识别出280个IMMAs,涵盖了约13%的已考海域。从比例上看,这些区域中,57%位于专属经济区(EEZs),另外43%位于公海地区。最近批准的33个IMMAs,位于东北大西洋和波罗的海,涵盖了丰富的海洋生物和关键的栖息地,如觅食和迁徙的座头鲸、蓝鲸和鳍鲸。
学而思思考题·举一反三
Q1、刚刚开完了的联合国《生物多样性公约》缔约方大会第十六次会议(CBD-COP16) 上,发布了《2024年保护地球报告》,对“昆蒙框架”目标3的首次全球进展官方评估数据显得不容乐观:目前,全球17.6%的陆地和内陆水域以及8.4%的海洋和沿海地区位于有记录的保护区和保护区内;要达到30*30的目标,那么,未来6年,全球保护覆盖的面积需要大大的拓展——陆地面积需再扩大12.4%,海洋面积需再扩大21.6%。
那么,在当前已识别的IMMAs数量有限的情况下,这是否反映了我们对全球海洋哺乳动物栖息地认知的不足?结合《昆明-蒙特利尔全球生物多样性框架》(GBF)提出的30*30目标,我们应如何系统性地加强对未识别海洋哺乳动物栖息地的研究与保护,以确保实现到2030年保护30%海洋和陆地生态系统的目标?
Q2、当前的保护措施能否有效应对气候变化带来的生态变化,尤其是在深潜鲸类栖息地的生境退化和食物链变化方面?我们应如何结合长期生态监测和模型预测,调整保护策略以应对这些潜在威胁?
Q3、在南海的海洋生物多样性保护中,如何有效整合鲸类及其栖息地的科学研究与当地渔业和经济活动的可持续发展,以实现生态保护与经济利益的双赢?
Q4、当前的保护措施能否有效应对气候变化带来的生态变化,尤其是在深潜鲸类栖息地的生境退化和食物链变化方面?我们应如何结合长期生态监测和模型预测,调整保护策略以应对这些潜在威胁?
Q5、在南海深潜鲸类的栖息地选择中,环境因素(如水温、盐度和水深)与生物因素(如猎物分布和竞争关系)如何交互作用,影响不同鲸类物种的种群动态和栖息地利用模式?
欢迎分享你的想法!(请文末留言)
THE END
海湿声明: 1.本文仅代表资讯,不代表平台观点。供参考。
2. 因本平台斜体字拷贝至外部平台时常出现内容丢失的情况,
故本文中,物种拉丁学名未作斜体设置。
编译 | 王芊佳
审核 | LI Songhai
编辑 | 绿茵
排版 | 绿叶
参考资料略


 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国